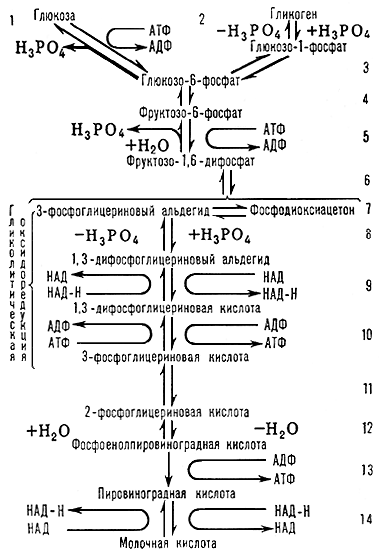
Гликолиз (от греч. glykys — сладкий и lysis — распад, разложение), процесс анаэробного ферментативного негидролитического расщепления углеводов (главным образом глюкозы) в животных тканях, сопровождающийся синтезом аденозинтрифосфорной кислоты (АТФ) (см. Аденозинфосфорные кислоты) заканчивающийся образованием молочной кислоты. Г. имеет большое значение для мышечных клеток, сперматозоидов, растущих (в т. ч. опухолевых) тканей, т.к. обеспечивает накопление энергии в отсутствии кислорода. Продукты, образующиеся при Г., являются субстратами последующих окислительных превращений (см. Трикарбоновых кислот цикл). Процессами, аналогичными Г., являются молочнокислое, маслянокислое, спиртовое и пр. виды брожения, протекающего в растительных, дрожжевых и бактериальных клетках. Интенсивность отдельных стадий Г. зависит от кислотности — водородного показателя — рН (оптимум рН 7—8), температуры и ионного состава среды. Последовательность реакций Г. (см. схему) хорошо изучена, идентифицированы промежуточные продукты, выделены ферменты Г. в кристаллическом или очищенном виде.
Г. начинается с образования фосфорных производных сахаров, что способствует превращению циклической формы субстрата в ациклическую, более реакционноспособную. Одной из реакций, регулирующих скорость Г., является реакция 2, катализируемая ферментом фосфорилазой. Существенную регуляторная роль принадлежит также ферменту фосфофруктокиназе (реакция 5), активность которой тормозится АТФ, но стимулируется продуктами её распада. Центральным звеном Г. является гликолитическая оксидоредукция (реакции 8—10), представляющая окислительно-восстановительный процесс, протекающий с окислением 3-фосфоглицеринового альдегида до 3-фосфоглицериновой кислоты и восстановлением кофермента никотинамидадениндинуклеотида (НАД). Эти превращения осуществляет дегидрогеназа 3-фосфоглицеринового альдегида (ДФГА) при участии фосфоглицераткиназы.
В результате оксидоредукции высвобождается энергия, аккумулирующаяся (в виде богатого энергией соединения — АТФ) в процессе субстратного фосфорилирования. Второй реакцией, обеспечивающей образование АТФ, является реакция 13. Г. кончается образованием молочной кислоты (реакция 14) под действием лактатдегидрогеназы и с участием восстановленного НАД. Т. о., при расщеплении 1 молекулы глюкозы образуются 2 молекулы молочной кислоты и 4 молекулы АТФ. В то же время на первых стадиях Г. (см. реакции 1, 5) затрачиваются 2 молекулы АТФ на 1 молекулу глюкозы. В процессе Г. выделяется только около 7% энергии, которая может быть получена при полном окислении глюкозы (до СО2 и Н2О).
Кроме глюкозы, в процесс Г. могут вовлекаться глицерин, некоторые аминокислоты и др. субстраты. В мышечной ткани, где основной субстрат Г. — гликоген, процесс начинается с реакций 2 и 3 и носит название гликогенолиза. Общим промежуточным продуктом для гликогенолиза и Г. является глюкозо-6-фосфат.
Все реакции Г. обратимы, кроме 1, 5 и 13. Однако можно получить глюкозу (реакция 1) или фруктозомонофосфат (реакция 5) из их фосфорных производных при гидролитическом отщеплении фосфорной кислоты в присутствии соответствующих ферментов; реакция 13 практически необратима, по-видимому, вследствие высокой энергии гидролиза фосфорной группировки (около 13 ккал/моль). Поэтому образование глюкозы из продуктов Г. идёт другим путём.
В присутствии O2 скорость Г. снижается (эффект Пастера). В некоторых тканях (например, опухолевые клетки, сетчатка, безъядерные эритроциты) возможен и интенсивный, т. н. аэробный, Г. в присутствии кислорода. Кроме того, имеются примеры подавления гликолизом тканевого дыхания (эффект Кребтри) в некоторых интенсивно гликолизирующих тканях. Механизмы взаимоотношений анаэробных и аэробных окислительных процессов до конца не изучены.
А. А. Болдырев.

Рис. к ст. Гликолиз.