Зародышевые листы
(или пласты, Keimblätter, feuillets blastodermiques, germinal layers) — термин, употребляемый в эмбриологии для обозначения тех первоначальных, основных клеточных зачатков, из которых образуется организм животного при его эмбриональном развитии. В статьях Дробление, Бластула и Гаструла были изложены те процессы, с которых начинается развитие многоклеточных животных и которые приводят к образованию двухслойного зачатка, состоящего из наружного слоя клеток — наружного листа, или эктодермы, и внутреннего — энтодермы (также epiblast и hypoblast). Главнейшие способы образования этих двух первичных пластов: впячивание, или инвагинация, обрастание, или эпиболия, миграция клеток и их отщепление, или деламинация. Впячивание в его наиболее простой форме наблюдается тогда, когда правильное дробление яйца приводит к образованию шара, поверхность которого состоит из одного слоя клеток (бластодерма), а полость выполнена жидкостью. Клетки бластодермы по своему расположению представляют собою слой эпителия. Одна половина этого шара, клетки которой отличаются обыкновенно несколько большей величиной и большим содержанием желтка, впячивается в его полость, вследствие чего образуется зародыш в виде чашечки или кубка — гаструла (таблица, рис. 1).
ЗАРОДЫШЕВЫЕ ЛИСТЫ И ОБОЛОЧКИ
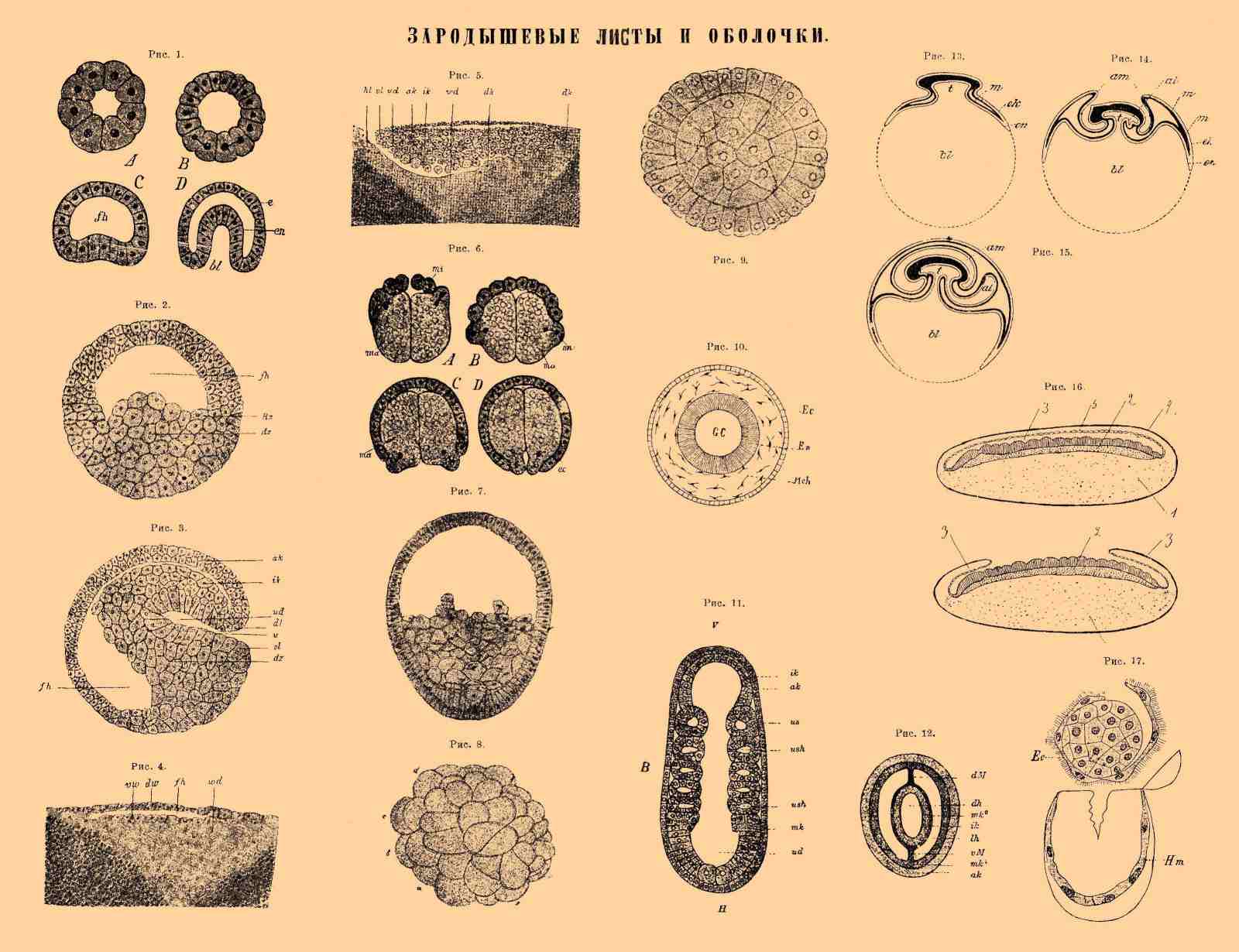
Рис. 1. Образование гаструлы путем выпячивания из бластулы; fh — полость дробления, e — эктодерма, en — энтодерма, bl — бластопор. Рис. 2. Бластула тритона: fn — полость дробления, dz — клетки, набитые желтком. Рис. 3. Разрез через яйцо тритона при начале гастрального втягивания: ak — наружный лист, ik — внутренний лист, fh — полость дробления, ud — archenteron, u -бластопор, dz — желточные клетки. Рис. 4. Разрез через зародышевую полоску цыпленка по окончании дробления (в стадии бластулы): fh — полость дробления, vw — нижняя, dw — верхняя стенка бластулы. Рис. 5. Гаструляция в зародышевой полости птиц: ak — наружный, ik — внутренний лист, dk — желточные ядра, ud — archenteron, hl, vl — губы бластопора. Рис. 6. Образование пластов путем эпиболии (в яйцах червя Bonellia): mi — мелкие клеточки (микромеры), дающие начало эктодерме (ec), ma — крупные клетки (макромеры), образующие энтодерму. Рис. 7. Образование энтодермы путем миграции клеток (в яйце медузы). Рис. 8. Образование пластов путем деламинации (яйцо медузы): morula. Рис. 9 Образование пластов путем деламинации (яйцо медузы): обособление двух пластов. Рис. 10. Схема строения пластов у животных с средним листом в виде мезенхимы: Ec — эктодерма, En — энтодерма, Gc — полость кишечника, Mch — мезенхима. Рис. 11. Зародыш ланцетника (Amphioxus) в оптическом продольном разрезе: V — передний, H — задний конец, ak — наружный, ik — внутренний лист, ud — полость кишки. Образование мезодермы путем отщепления парных (целомических) мешочков (us) от энтодермы: ush — целомическая полость. Рис. 12. Схематический поперечный разрез через животное с двумя мезодермическими листками и вторичною полостью тела: ak — эктодерма, ik — энтодерма, dh — полость кишки. Средний лист (мезодерма) образуется из двух мешочков, прилегающих друг к другу на спинной и брюшной стороне для образования спинного (dM) и брюшного (vM) мезентерия. Каждый мешочек одной стенки прилегает к кишке, образуя висцеральный (кишечный) лист мезодермы, mk2, другою — в коже, образуя париетальный (кожный) листок mk1. В промежутке между обоими листками образуется вторичная (целомическая) полость тела. Рис. 13-15. Последовательные стадии образования зародышевых оболочек у птицы: bl — питательный желток, t — полость кишечника, ek — эктодерма, en — энтодерма, m — мезодерма, am (Рис. 14) — складки, из которых образуются зародышевые оболочки: am (Рис. 15) — амнион, s — сероза, al — аллантоис. 16. Образование зародышевых оболочек у насекомых: 1 — питательный желток, 2 — зародышевая полоска, 3 — эктодермические складки вокруг зародышевой полоски, своим разрастанием образующие оболочки, 4 — наружная зародышевая оболочка, serosa, непосредственно переходящая в эктодерму, покрывающую все яйцо, 5 — внутренняя зародышевая оболочка, amnion, 6 — полость между зародышем и amnion. 17. Зародыш глисты (лентеца широкого Bothricephalus latus), выдавленный из яйца. Ec — эктодерм, Hm — зародышевая оболочка.
Стенка этого кубка состоит из двойного ряда клеток — наружный представляет собой эктодерму, внутренний, выстилающий первичную пищеварительную полость гаструлы (archenteron) — энтодерму; отверстие, при посредстве которого полость archenteron'a открывается наружу, наз. бластопор. Такой способ образования двух первичных зародышевых листов, являющихся в виде эпителиальных пластинок, наблюдается у иглокожих, ланцетника (Amphioxus), у червя Sagitta и др. Но образование пластов путем впячивания может происходить и при других способах дробления, подвергаясь различным изменениям в зависимости, главным образом, от количества питательного желтка, заключающегося в яйце. Так, напр., у речного рака (Astacus) стадия бластулы выражена в виде однослойного бластодермического пузыря, полость которого выполнена питательным желтком. Впячивание происходит на весьма ограниченной части поверхности (соответствующей будущей брюшной стороне животного). Впячивающиеся клетки, представляющие собою энтодерму, сперва вдаются в желток; затем они начинают постепенно поглощать желток, вбирая его в себя и вырастая таким путем в высокие, пирамидальные, набитые желтком клетки; ядра клеток, окруженные протоплазмой, сохраняют при этом свое положение у основания энтодермического эпителия. Когда клетки энтодермы пожрут весь желток, так что доходят своими основаниями до противоположной стенки бластодермы, то они вновь освобождаются от желтка, выделяя его уже в пищеварительную полость: желток перемещается таким образом из полости дробления (blastocoel) в архентерическую полость, откуда он мало-помалу и усваивается зародышем. Образование двух первичных пластов у позвоночных также может быть сведено к образованию инвагинационной гаструлы. Всего проще процесс гаструляции у земноводных, напр. у тритона, у саламандры (табл., рис. 2 и 3). Здесь продуктом неравномерного дробления является толстостенная бластула, в которой можно различить два полюса: на одном стенка более тонка и состоит из нескольких рядов мелких клеточек; на противоположном, вегетативном полюсе значительно утолщенная стенка состоит из крупных, набитых желтком клеток, выдающихся бугром в blastocoel, который оттесняется ими к другому полюсу и лежит, следов., эксцентрически. Впячивание начинается на одной стороне яйца в виде узкой, резко обозначенной борозды, появляющейся на границе между верхними, мелкими, и нижними, крупными, клетками. При дальнейшем впячивании от верхнего края борозды впячиваются в глубину яйца мелкие клетки верхней половины, от нижнего — крупные клетки нижней половины яйца; первые образуют как бы крышку, вторые — как бы дно архентерона. Последний сначала имеет вид лишь узкой щели рядом с широкой полостью дробления; позднее же он ее совсем вытесняет и расширяется на дне впячивания в широкий мешок, суживающийся к бластопору. Когда впячивание закончено, все клетки нижней, вегетативной стороны яйца уходят внутрь гаструлы, выстилая архентерон; по поверхности яйца распределяются тогда в один слой (у лягушки в 2-3 слоя) клетки верхней половины. Так. обр. у амфибий впячивается внутрь вся толстая масса желточных клеток; то же самое происходит при развитии яйца у круглоротых рыб (Суclostomata). У остальных рыб, у пресмыкающихся и птиц вследствие огромного скопления питательного желтка дробление сделалось частичным (меробластические яйца), и инвагинационная гаструла является, так сказать, в искаженном виде (табл., рис. 4 и 5). Полость дробления сводится в их яйцах к узкой щели; лишь верхняя стенка, кровля этой щели, состоит из клеток, образующих зародышевый кружок; дно щели, нижнюю стенку ее, составляет нераздробившийся желток, заключающий в себе лишь по соседству с зародышевым кружком отдельные желточные ядра. Процесс впячивания ограничивается зародышевым кружком: не подвергающийся дроблению желток уже по одному своему объему не может впячиваться. Начало впячивания полагается появлением на одной стороне зародышевого кружка серповидной бороздки, отделяющей кружок от подлежащего желтка. От этой бороздки начинается процесс разрастания клеточек, образующих нижний, внутренний слой зародышевого кружка и превращающих его в двухслойный зачаток. Архентерическая полость является в форме узкого щелевидного промежутка, ограниченного лишь сверху клетками, снизу же — нераздробившимся желтком. Об образовании пластов у позвоночных см. Hertwig, "Lehrbuch der Entwickelungsgeschichte des Menschen u. der W i rbelthiere" (Иена, 1893).
Во всяком случае у позвоночных, как бы велико ни было скопление питательного желтка, всегда можно найти при образовании З. пластов более или менее ясные следы инвагинационного процесса. Но в других случаях, именно, у многих беспозвоночных (ктенофор, аннелид) после неравномерного дробления яиц, богатых питательным желтком, З. листы образуются путем обрастания (эпиболии, рис. 6 табл.). В таких яйцах по окончании дробления на одном полюсе яйца лежат мелкие, лишенные желтка клетки, а на другом — крупные, богатые желтком. Последние и представляют собою энтодерму; но впячивание их оказывается невозможным вследствие малого развития или даже полного отсутствия полости дробления. Мелкие клеточки, лежащие на одном полюсе яйца, размножаясь в сторону вегетативного полюса, мало-помалу обрастают всю плотную массу энтодермических клеток. В результате получается двухслойный зачаток, где слой эктодермических клеток покрывает собою плотную массу клеток энтодермы, не выстилающих, однако, никакой полости: в такой гаструле нет архентерона. Лишь гораздо позднее, под конец развития, между клетками энтодермы начинает появляться полость кишечника. Границу эктодермы при постепенном обрастании ею энтодермы можно рассматривать как бластопор. Способ миграции (табл., рис. 7) заключается в том, что в бластуле, заключающей в себе обширную сегментационную полость, для образования энтодермы известные клетки бластодермы или сами уходят с поверхности в глубину полости дробления, или же путем деления отщепляют от себя клетки, которые и мигрируют в названную полость. Такое выхождение клеток с поверхности в глубину яйца может происходить по всей его поверхности или же быть сосредоточенным на одном полюсе. В результате получается двухслойный зачаток с поверхностным слоем эктодермы и с плотною массою энтодермических клеток, в среде которых пищеварительная полость обособляется лишь позднее. Такой способ образования пластов обыкновенен среди кишечнополостных животных (Coelenterata), но к нему же очень подходит и образование пластов y некоторых суставчатоногих животных, напр. у Peripatus или у пауков (Araneidae). Образование пластов путем деламинации может совершаться двумя способами: 1) путем поперечного деления всех клеток бластодермы, которая целиком распадается при этом на два эпителиальных слоя, из которых внутренний представляет собою энтодерму; 2) путем обособления поверхностных клеток плотного многоклеточного зачатка, образовавшегося после дробления яйца (такое плотное скопление клеток дробления, без сегментационной полости, носит название morula), в эктодерму, причем все остающиеся центральные бластомеры представляют собою энтодерму (табл., рис. 8 и 9). Оба последние способа образования З. пластов (путем миграции клеток и путем деламинации) изучены Мечниковым ("Embryologische Studien an Medusen", Вена, 1886). Деламинация встречается почти исключительно между кишечнополостными; у остальных животных она наблюдалась лишь в очень редких случаях. Так, деламинацию описывают у пауков-сенокосцев (Фаусек), у рака — Limulus (Klngsteу).
До сих пор мы говорили об образовании только двух первичных З. пластов — экто— и энтодермы. У низших представителей многоклеточных животных, у кишечнополостных (Coelenterata), дифференцирование эмбриональных элементов в некоторых случаях далее этого и не идет. Все тело гидры состоит из двух эпителиальных пластинок — одной, составляющей ее наружный покров, и другой, выстилающей пищеварительную полость. При эмбриональном развитии этого животного эктодерма и энтодерма зародыша непосредственно превращаются в два соответственных эпителиальных слоя окончательной формы. Но у части кишечнополостных и у всех вышестоящих Metazoa при эмбриональном развитии между экто— и энтодермой появляется еще особый клеточный зачаток — группа эмбриональных клеток, составляющая средний пласт, или мезодерму (мезобласт). Из этих трех З. листов развиваются всегда вполне определенные, равнозначащие системы органов. Так, из эктодермы образуется: 1) эпителиальный покров тела со множеством его производных: различного рода железами, кутикулярными оболочками, хитином, волосами, роговыми образованиями и т. д.; 2) нервная система; 3) важнейшие в функциональном отношении части органов чувств (чувствительные клетки). Из клеток энтодермы развивается эпителиальная выстилка известной части пищеварительного канала и пищеварительных желез (передняя и задняя части пищеварительного канала часто образуются путем впячивания эктодермы). К производным среднего пласта относятся некоторые железы (орг. выделения и обыкновенно половые железы), мускулатура и все элементы опоры — соединительная ткань с ее производными.
Однако мезодерма, или средний пласт, ни в каком случае не может быть поставлена в ряд с двумя первыми пластами — уже по тому одному, что в огромном большинстве случаев она составляет производное одного из первичных З. пластов, всего чаще — энтодермы. Уже одно это указывает на ее подчиненное положение по отношению к первичным пластам. Касательно двух первичных З. пластов между учеными не существует разногласий как в их нахождении у всех Metazoa, так и относительно того, что повсюду у всех животных эти два пласта представляют, несмотря на различия в их способе развития, образования вполне соответствующие друг другу, гомологичные. Способы же развития мезодермы, ее отношения к двум первичным пластам представляют столько фундаментальных различий, что распространять гомологию между зародышевыми пластами всех животных и на все те разнообразные образования, которые обозначаются именем среднего пласта, кажется вряд ли возможным. Раздаются даже голоса, утверждающие, что мезодермы как самостоятельного, обособленного зачатка известной суммы органов вообще не существует и что все те образования, которым приписывается обыкновенно мезодермическое происхождение, должны быть отнесены непосредственно к производным одного из двух первичных пластов (Kleinenberg). Остановимся лишь на нескольких примерах образования среднего пласта у различных животных.
Различают мезенхиму от мезодермы собственно. Мезенхима образуется путем выхождения, миграции, отдельных клеточек из бластодермического эпителия в сегментационную полость; такие клетки, освободившись от связи с другими, принимают амебовидную форму и вместе с этим известную самостоятельность; образуется ткань, состоящая из отдельных разбросанных, свободных клеток, наполняющих промежуток между экто— и энтодермой (табл., рис. 10). Мезодерма собственно является в виде симметрично расположенных парных клеточных полосок или мешочков, стенки которых принимают эпителиальный характер. Мешочки эти расположены по обе стороны энтодермического зачатка кишечника, между ним и эктодермой. У членистых животных (кольчатые черви, суставчатоногие, отчасти также позвоночные) мешочки эти вытянуты попарно в ряд по продольной оси зародыша, образуя мезодермические сомиты. Полость таких мешочков называется целомическою (coelom) и дает начало будущей так назыв. вторичной полости тела, выстланной слоем так назыв. перитонеального эпителия. Одна стенка каждого такого целомического мешочка прилегает к эктодерме, образуя соматический, или паретальный, лист мезодермы; другая в виде спланхнического, или висцерального, листа, прилегает к энтодерме. На спинной и брюшной стороне зародыша вдоль срединной линии мезодермические мешочки каждой стороны сходятся между собою, образуя подвески кишечника, так наз. мезентерии (табл., рис. 12). Такой характер носит мезодерма у кольчатых червей (Annelides), строение которых можно принять за исходную точку при изучении развития среднего листа и его производных у остальных Metazoa. Между низшими типами беспозвоночных, у губок, кишечнополостных, плоских червей — мезодерма выражена исключительно в виде более или менее сильно развитой мезенхимы; у остальных типов существуют обыкновенно одновременно и мезенхима, и распадающаяся на два слоя для образования целомической полости мезодерма. У моллюсков мощно развитая мезенхима оттесняет на задний план мезодерму с ее целомической полостью; у некоторых червей (Sagitta), напротив, мезенхима почти исчезает. В эмбриональном развитии позвоночных появляется как мезодерма, распадающаяся на сегменты и на два листка, париетальный и висцеральный, так и мезенхима; последняя образуется путем выхождения эпителиальных клеток из З. листков, клеток, проникающих в виде блуждающих элементов в промежуток между четырьмя листками (остаток первоначальной полости дробления). Этим мезенхиматическим элементам приписывают образование крови и соединительной ткани.
Обособление мезодермы от первичных пластов происходит у различных животных неодинаково. У кольчатых червей (Annelides) на краю бластопора обособляются по обе стороны от срединной плоскости по одной крупной клетке энтодермы, которые вклиниваются затем между двумя первичными листами. Это так назыв. образующие клетки мезодермы (Urmesodermzellen). Путем размножения этих клеток образуются две клеточные полоски (мезодермические полоски), распадающиеся затем на описанные выше парные, заключающие полость внутри, целомические мешочки. От тех же крупных образующих клеток мезодермы берут начало и элементы мезенхимы. Подобные же образующие клетки (Urmesodermzellen) для среднего листа (мезодермы и мезенхимы) наблюдаются у моллюсков. Другой способ образования мезодермы наблюдается в особенно чистом виде у червя Sagitta. Здесь целомические мешочки образуются в виде складок энтодермы, в виде выпячивания стенок кишечника гаструлы (ar c henteron). Целомическая полость стоит первоначально в непосредственной связи с полостью архентерона и составляет отделившуюся ее часть. Подобное же образование мезодермы и вторичной полости тела мы видим у брахиопод, у иглокожих. Между позвоночными оно в резко выраженной форме наблюдается у ланцетника, Amphioxus, где появляются два ряда попарно отщепляющихся от полости архентерона целомических мешочков (табл., рис. 11). Образование мезодермы у остальных позвоночных представляет собой также более или менее явственное видозмененение того же способа.
История учения о З. листах есть в то же время история всей сравнительной эмбриологии. Уже основатель эмбриологии животных К. Ф. Вольф в половине прошлого столетия наблюдал, что кишечный канал цыпленка залагается в виде листа, загибающегося потом в желобок и наконец замыкающегося в трубку. Подобный же способ развития приписывал он и другим системам органов. Пандер в 1817 г. различал в 12 часовом зародыше цыпленка две тонкие, легко отделяющиеся друг от друга пластинки, между которыми позднее появляется еще третья. Настоящим основателем учения о З. листах является К. Э. ф. Бэр, изложивший свои многолетние наблюдения в знаменитом труде "Ueber Entwickelungsgeschichte d. Thiere. Beobachtung u. Reflexion". Ремах и Келлинер первые применили к З. листам учение о клеточном строении организма. Первоначально учение о З. пластах было выработано лишь по отношению к эмбриологии позвоночных. Английский натуралист Гексли в 1849 г. первый провел параллель между двумя клеточными перепонками, из которых состоит тело кишечнополостных, и двумя первичными листками зародыша позвоночных. Исследования русских ученых, Ковалевского и Мечникова, произведенные в конце 60-х годов, показали применимость учения о З. листах к эмбриональному развитию всех вообще многоклеточных животных. Основываясь на многочисленных наблюдениях Ковалевского, Геккель построил свою теорию гаструлы как зародышевой формы, состоящей из двух листков — экто— и энтодермы — и представляющей собой исходную точку в развитии всех животных, кроме простейших. В учении о развитии мезодермы и полости тела сыграли важную роль также наблюдения Ковалевского и Мечникова над развитием иглокожих, брахиопод, Sagitta и Amphioxus. Из сочинений, заключающих в себе теоретическое обсуждение разных сторон вопроса о З. пластах, укажем еще на работу Мечникова "Embryologische Studien an Medusen", o которой мы уже говорили выше; в ней автор, основываясь на изучении эмбриологии низших современных Metazoa, делает веские возражения против Геккелевской теории гаструлы. Ср. книгу братьев Гертвигов "Die Coelomtheorie" (Иена, 1881); Kleinenberg, "Die Entstehung d. Annelids aus der Larve von Lopadorhynchus" ("Zeit. wiss. Zool.", 44 т. 1886). Литературу о З. листах см. в вышеназванном учебнике эмбриологии позвоночных Гертвига. См. также Богданов, "Медицинская зоология", т. II.
В. Фаусек.