Радиочувствительность, чувствительность биологических объектов к действию ионизирующих излучений. Облучение вызывает в клетках и организмах различные изменения (см. Биологическое действие ионизирующих излучений),степень проявления которых не всегда коррелирует между собой. Поэтому при оценке Р. важно учитывать, какой критерий используется для её характеристики. Обычно таким критерием служит летальное действие излучений — инактивация или гибель клеток и гибель многоклеточных организмов. Летальное действие излучении также может проявляться в разных формах: в случае клеток — гибель их в интерфазе после одного или нескольких делений (см. Митоз),в случае многоклеточных организмов — гибель в разные сроки после облучения.
Чтобы оценить Р., биологические объекты облучают разными дозами, определяют процент выживших и строят кривые выживания. Для клеток такие кривые изображают обычно в полулогарифмическом масштабе (рис. 1), для многоклеточных организмов — в линейном (рис. 2). Пользуясь кривыми выживания, находят ЛД50 — дозу, после которой выживает 50% особей, а также значения DQ и D0, отражающие величину «плеча» и наклон прямолинейной составляющей таких кривых (значение D0 равно дозе, уменьшающей выживаемость в е 2,7 раза на прямолинейной составляющей кривой выживания). В экспериментах с млекопитающими ЛД50 определяют обычно для разных сроков после облучения — 3, 5, 15, 30 и т.д. суток. Получаемые значения ЛД50/5, ЛД50/30 и т.п. отражают Р. тех систем организма, преимущественное поражение которых ответственно за его гибель в течение того или иного отрезка времени. Так, гибель мышей и крыс в течение первых 3—5 сут после облучения связана с повреждением кишечного тракта, а в интервале между 5 и 30 сут — с повреждением системы кроветворения. Мерой Р. обычно служат ЛД50 или D0.
Р. клеток может различаться в сотни и тысячи раз: ЛД50 для клеток млекопитающих — 200—350 рад, для бактерий и дрожжей — 10—45 тыс. рад, для инфузорий и амёб — 300—500 тыс. рад. Р. обусловливается первичной поражаемостью жизненно важных структур клеток, их способностью к восстановлению (репарации) и условиями культивирования. В общем случае Р. клеток растет с увеличением содержания ДНК, числа и размеров хромосом и уменьшается с увеличением числа хромосомных наборов (плоидности). Вместе с тем на Р. клеток влияют их химический состав (например, содержание эндогенных тиолов), физиологическое состояние (фаза клеточного цикла, фаза дифференцировки), условия во время облучения (могут оказывать радиозащитное или радиосенсибилизирующее действие) и условия в пострадиационный период (могут способствовать или препятствовать осуществлению репарации и проявлению первичных повреждений). Клетки с нарушенной системой репарации отличаются повышенной Р. Мутации в отдельных генах могут в десятки раз изменять Р. клеток, влияя на различные стороны метаболизма. Т. о., Р. клеток зависит от многих факторов, удельный вес которых у разных объектов различен. Р. многоклеточных растений и животных также широко варьирует. Так, для семян гороха и кукурузы ЛД50 равна 5—20 тыс. рад, для семян клевера и редиса — 100—250 тыс. рад (для проростков этих же растений ЛД50 составляет 250—700 рад); для взрослых насекомых ЛД50 — 30—50 тыс. рад, а для млекопитающих — от 350—700 до 1000—1200 рад. Р. растений и животных обусловливается главным образом Р. их клеток (в случае млекопитающих — Р. стволовых клетоких кроветворных органов и желудочно-кишечного тракта) и факторами, влияющими на успешность регенерации поврежденных облучением органов и тканей за счёт размножения выживших клеток. На проявление Р. влияют условия содержания после облучения, способствующие или препятствующие выздоровлению от лучевой болезни. Помимо биологических особенностей и условий среды, Р. клеток и организмов зависит от физических свойств излучений, мощности дозы и особенностей фракционирования облучения. Разработаны способы радиосенсибилизации, т. е. искусственного увеличения Р. биологических объектов. Изучение различных аспектов Р. важно для разработки эффективных методов лечения лучевых повреждений, радиотерапии раковых опухолей, а также в случаях применения излучений для радиостимуляции растений и в искусственном мутагенезе.
Лит.: Основы радиационной биологии, М., 1964; Тимофеев-Ресовский Н. В., Иванов В. И., Корогодин В. И., Применение принципа попадания в радиобиологии, М., 1968; Кузин А. М., Структурно-метаболическая гипотеза в радиобиологии, М., 1970; Акоев И. Г., Максимов Г. К., Малышев В. М., Лучевое поражение млекопитающих и статистическое моделирование, М., 1972; Мясник М. Н., Генетический контроль радиочувствительности бактерий, М., 1974.
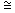 2,7 раза на прямолинейной составляющей кривой выживания). В экспериментах с млекопитающими ЛД50 определяют обычно для разных сроков после облучения — 3, 5, 15, 30 и т.д. суток. Получаемые значения ЛД50/5, ЛД50/30 и т.п. отражают Р. тех систем организма, преимущественное поражение которых ответственно за его гибель в течение того или иного отрезка времени. Так, гибель мышей и крыс в течение первых 3—5 сут после облучения связана с повреждением кишечного тракта, а в интервале между 5 и 30 сут — с повреждением системы кроветворения. Мерой Р. обычно служат ЛД50 или D0.
2,7 раза на прямолинейной составляющей кривой выживания). В экспериментах с млекопитающими ЛД50 определяют обычно для разных сроков после облучения — 3, 5, 15, 30 и т.д. суток. Получаемые значения ЛД50/5, ЛД50/30 и т.п. отражают Р. тех систем организма, преимущественное поражение которых ответственно за его гибель в течение того или иного отрезка времени. Так, гибель мышей и крыс в течение первых 3—5 сут после облучения связана с повреждением кишечного тракта, а в интервале между 5 и 30 сут — с повреждением системы кроветворения. Мерой Р. обычно служат ЛД50 или D0.
