Рибосомы, внутриклеточные частицы, осуществляющие биосинтез белка; Р. обнаружены в клетках всех без исключения живых организмов: бактерий, растений и животных; каждая клетка содержит тысячи или десятки тысяч Р.
Форма Р. близка к сферической, хотя её очертания сложны и не могут быть описаны простой геометрической фигурой. Различают 2 главных класса Р.: так называемые 70 SP (молекулярная масса около 3×106, диаметром около 200—300 , коэффициент седиментации S°20w около 70 единиц Сведберга) и более крупные 80 S P. (молекулярная масса около 4—5×106, максимальный размер до 400 , коэффициент седиментации около 80 единиц Сведберга). Р. 70 S класса характерны для клеток, не имеющих оформленного ядра, — прокариотов (бактерии, актиномицеты и синезелёные водоросли), а также для хлоропластов и митохондрийвысших организмов. Р. 80 S класса обнаружены в цитоплазме всех эукариотов, т. е. организмов, имеющих оформленное клеточное ядро. По химической природе Р. — нуклеопротеид, состоящий из рибонуклеиновой кислоты (РНК) и белка. Р. класса 70 S содержит 60—65% РНК и 40—35% белка, а Р. класса 80 S — около 50% РНК и 50% белка. Универсальный принцип структурной организации Р. — построение её из двух неравных субчастиц (субъединиц), на которые она может диссоциировать (например, при понижении концентрации ионов Mg2+ в среде) и вновь реассоциировать по схеме:
70 S Û 50 S + 30 S; 80 S Û 60 S + 40 S
Большая субчастица (50 S или 60 S) состоит из молекулы высокополимерной рибосомальной РНК (молекулярная масса 1,1—1,8×106), молекулы относительно низкополимерной рибосомальной РНК (молекулярная масса 40 000) и нескольких десятков молекул белков. Малая субчастица (30 S или 40 S) состоит из молекулы высокополимерной рибосомальной РНК (молекулярная масса 0,6—0,7×106) и от 20 (в 30 S частицах) до 40 (в 40 S частицах) различных молекул белков. Высокополимерная рибосомальная РНК создаёт возможность сборки этих белков в единую рибонуклеопротеидную частицу. В эксперименте можно осуществить разворачивание Р.: частица становится более рыхлой, РНК разворачивается в тяж, при этом все белки остаются связанными с ней. В других условиях можно добиться последовательного отделения белков от РНК (разборка Р.). Эта разборка обратима, и в подходящих условиях белки и РНК снова спонтанно объединяются в рибонуклеопротеид, формирующий нативную структуру Р. (самосборка Р.). Считают, что образование Р. в клетках также идёт путём самосборки из предварительно синтезированных РНК и белков.
В процессе функционирования (т. е. синтеза белка) Р. осуществляет несколько функций: 1) специфическое связывание и удержание компонентов белоксинтезирующей системы [информационная, или матричная, РНК (иРНК): аминоацил-тРНК; пептидил-тРНК; гуанозинтрифосфат (ГТФ); белковые факторы трансляции EF — Т и EF — G]: 2) каталитические функции (образование пептидной связи, гидролиз ГТФ): 3) функции механического перемещения субстратов (иРНК, тРНК), или транслокации. Функции связывания (удержания) компонентов и катализа распределены между двумя рибосомными субчастицами. Малая рибосомная субчастица содержит участки для связывания иРНК и аминоацил-тРНК и, по-видимому, не несёт каталитических функций. Большая субчастица содержит каталитический участок для синтеза пептидной связи, а также центр, участвующий в гидролизе ГТФ: кроме того, в процессе биосинтеза белка она удерживает на себе растущую цепь белка в виде пептидил-тРНК. Каждая из субъединиц может проявить связанные с ней функции отдельно, без связи с другой субчастицей. Однако ни одна из субчастиц в отдельности не обладает функцией транслокации, осуществляемой только полной Р. О функционировании Р. см. в статьях Белки, Трансляция.
Лит.: Спирин А. С., Гаврилова Л. П., Рибосома, 2 изд., М., 1971.
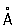 , коэффициент седиментации S°20w около 70 единиц Сведберга) и более крупные 80 S P. (молекулярная масса около 4—5×106, максимальный размер до 400
, коэффициент седиментации S°20w около 70 единиц Сведберга) и более крупные 80 S P. (молекулярная масса около 4—5×106, максимальный размер до 400  , коэффициент седиментации около 80 единиц Сведберга). Р. 70 S класса характерны для клеток, не имеющих оформленного ядра, — прокариотов (бактерии, актиномицеты и синезелёные водоросли), а также для хлоропластов и митохондрий высших организмов. Р. 80 S класса обнаружены в цитоплазме всех эукариотов, т. е. организмов, имеющих оформленное клеточное ядро. По химической природе Р. — нуклеопротеид, состоящий из рибонуклеиновой кислоты (РНК) и белка. Р. класса 70 S содержит 60—65% РНК и 40—35% белка, а Р. класса 80 S — около 50% РНК и 50% белка. Универсальный принцип структурной организации Р. — построение её из двух неравных субчастиц (субъединиц), на которые она может диссоциировать (например, при понижении концентрации ионов Mg2+ в среде) и вновь реассоциировать по схеме:
, коэффициент седиментации около 80 единиц Сведберга). Р. 70 S класса характерны для клеток, не имеющих оформленного ядра, — прокариотов (бактерии, актиномицеты и синезелёные водоросли), а также для хлоропластов и митохондрий высших организмов. Р. 80 S класса обнаружены в цитоплазме всех эукариотов, т. е. организмов, имеющих оформленное клеточное ядро. По химической природе Р. — нуклеопротеид, состоящий из рибонуклеиновой кислоты (РНК) и белка. Р. класса 70 S содержит 60—65% РНК и 40—35% белка, а Р. класса 80 S — около 50% РНК и 50% белка. Универсальный принцип структурной организации Р. — построение её из двух неравных субчастиц (субъединиц), на которые она может диссоциировать (например, при понижении концентрации ионов Mg2+ в среде) и вновь реассоциировать по схеме: