Спорообразование, спорогенез, процесс образования спор. У растительных организмов — прокариотов, клетки которых не имеют типичных ядер, споры могут возникать: из целой клетки, накопившей питательные вещества и утолщившей оболочку (экзоспоры многих сине-зеленых водорослей); при делении протопласта на большое число спор (эндоспоры некоторых сине-зеленых водорослей, рис. 1, 1); в результате уплотнения и сжатия протопласта внутри оболочки клетки и образования поверх него новой многослойной оболочки (у бактерий); при распаде особых участков мицелия на членики (у актиномицетов, рис. 1, 2). У растений — эукариотов, обладающих типичными ядрами, имеющих 3 основных типа спор (оо-, мито- и мейоспоры) и занимающих разное место в циклах развития, могут быть соответственно и 3 варианта С.: ооспорогенез, митоспорогенез и мейоспорогенез. Обычно под С. понимают образование мейоспор (мейоспорогенез). Ооспорогенез связан с процессом оплодотворения и, следовательно, со сменой ядерных фаз в циклах развития; заканчивается образованием ооспор (у многих зелёных водорослей и оомицетов), ауксоспор (у диатомей), зигоспор (у зигомицетов), представляющих собой одноядерные или многоядерные зиготы. Митоспорогенез приводит к возникновению митоспор, формирующихся по нескольку или в обльшом числе в результате митотических делений (см. Митоз) гаплоидных [например, зооспоры ряда водорослей (рис. 1, 3) и грибов], реже диплоидных (например, карпоспоры большинства флоридей) клеток или без делений — моноспоры эдогониума (рис. 1, 4), бангиевых, немалионовых; к смене ядерных фаз не приводит. Протекает в одноклеточных митоспорангиях (например, в зооспорангиях улотрикса, моноспорангиях эдогониума, цистокарпиях флоридей), а одноклеточные водоросли как бы сами становятся спорангиями (рис. 1, 5). Митоспорогенез может наблюдаться при распадении мицелия, состоящего из клеток, содержащих дикарионы, например у головнёвых и ржавчинных грибов. Мейоспорогенез связан со сменой диплофазы в циклах развития как низших, так и высших растений гаплофазой. У низших растений мейоспоры возникают в результате мейоза или вскоре после него из митотически разделившихся гаплоидных клеток, образовавшихся при мейозе. У водорослей и грибов с гаплоидным циклом развития С. происходит при прорастании зиготы (ооспоры), диплоидное ядро которой, делясь мейотически, образует 4 гаплоидных ядра; при этом возникают 4 мейоспоры (например, зооспоры хламидомонад, рис. 1, 6, апланоспоры улотрикса), либо 3 из четырёх гаплоидных ядер отмирают и образуется лишь 1 мейоспора (например, у спирогиры, рис. 1, 7), либо за мейозом следует 1—3 митотических деления и формируются 8—32 споры (например, у бангиевых). У водорослей, имеющих изоморфный и гетероморфный циклы развития, мейоспорогенез протекает в одноклеточных мейоспорангиях и характеризуется образованием либо 4 мейоспор (например, тетраспоры бурых водорослей и большинства флоридей, рис. 1, 8), либо 16—128 мейоспор (например, зооспоры ламинариевых, рис. 1, 9) вследствие 2—5 митотических делений, следующих за мейозом. В спорангиях сумчатых грибов (сумках, или асках) возникшие в результате мейоза 4 гаплоидных ядра делятся митотически и формируются 8 эндогенных мейоспор (аскоспор). В базидиях (спороносных органах) базидиальных грибов после мейоза возникают по 4 гаплоидных ядра, которые перемещаются в специальные выросты на поверхности базидий; в дальнейшем эти выросты с гаплоидными ядрами, т. и. базидиоспоры, отделяются от базидий (рис. 1, 10). Высшие растения образуют только мейоспоры, мейоспорогенез протекает в многоклеточных спорангиях. Обычно в результате митотических делений диплоидных клеток археспория возникают т. н. спороциты (мейотически делящиеся клетки), формирующие по 4 споры (тетрады спор). Равноспоровые папоротникообразные продуцируют морфологически и физиологически одинаковые споры (рис. 2, 1), из которых развиваются обоеполые заростки. У разноспоровых папоротникообразных и семенных растений осуществляются микро- и мегаспорогенез, мейоспорогенез, т. е. возникают споры двух типов. Микроспорогенез происходит в микроспорангиях и завершается образованием большого числа микроспор (рис. 2, 2), прорастающих затем в мужские заростки; мегаспорогенез — в мегаспорангиях, где в меньшем числе — часто даже 4 или 1 — созревают мегаспоры (рис. 2, 3), прорастающие в женские заростки. Развивающиеся спороциты и споры (у большинства высших растений) питаются веществами, получаемыми из клеток тапетума (слоя, выстилающего изнутри полость спорангия). У многих растений клетки этого слоя, расплываясь, образуют периплазмодий (протоплазматическую массу с дегенерирующими ядрами), в котором оказываются спороциты, а затем и споры. У некоторых растений в формировании периплазмодия участвует и часть спороцитов. В мегаспорангиях (семезачатках) некоторых покрытосеменных в результате мейоза образуются клетки с 2 или 4 гаплоидными ядрами, соответствующие 2 (рис. 2, 4) или 4 (рис. 2, 5) мегаспорам; из этих клеток развиваются женские гаметофиты — т. н. биспорические и тетраспорические зародышевые мешки. О С. у простейших см. в ст. Споры.
Лит.: Мейер К. И., Размножение растений, М., 1937; Курсанов Л.И., Комарницкий Н. А., Курс низших растений, М., 1945; Магешвари П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Тахтаджян А. Л., Высшие растения, т. 1, М. — Л., 1956; Поддубная-Арнольди В,А., Общая эмбриология покрытосеменных растений, М., 1964: Smith G. М., Cryptogamic botany, 2 ed., v. 1—2, N. Y. — L., 1955; Lehrbuch der Botanik für Hochschulen, 29 Aufl., Jena, 1967.
А. Н. Сладков.

Рис. 2. Спорообразование у высших растений. 1 — развитие спорангия у равноспорового лептоспорангиатного папоротника; 2 — развитие микроспорангия у селагинеллы (Selaginella); 3 — развитие микроспорангия у азолии (Azolla); 4 — мегаспороцит (а) до мейоза и возникшие из него клетки после первого (б) и второго (в) делений мейоза у лука (Allium cepa); 5 — мегаспороцит (а) до мейоза и образовавшиеся после первого и второго делений мейоза двуядерная (б) и четырехъядерные (в, г) клетки у рябчика (Fritillaria persica). АС — археспорий, ТП — тапетум, ПП — периплазмодий, СЦ — спороциты, ЯС — ядра спор, СП — споры (изоспоры), МС — мегаспора, ТМ — тетрады микроспор.
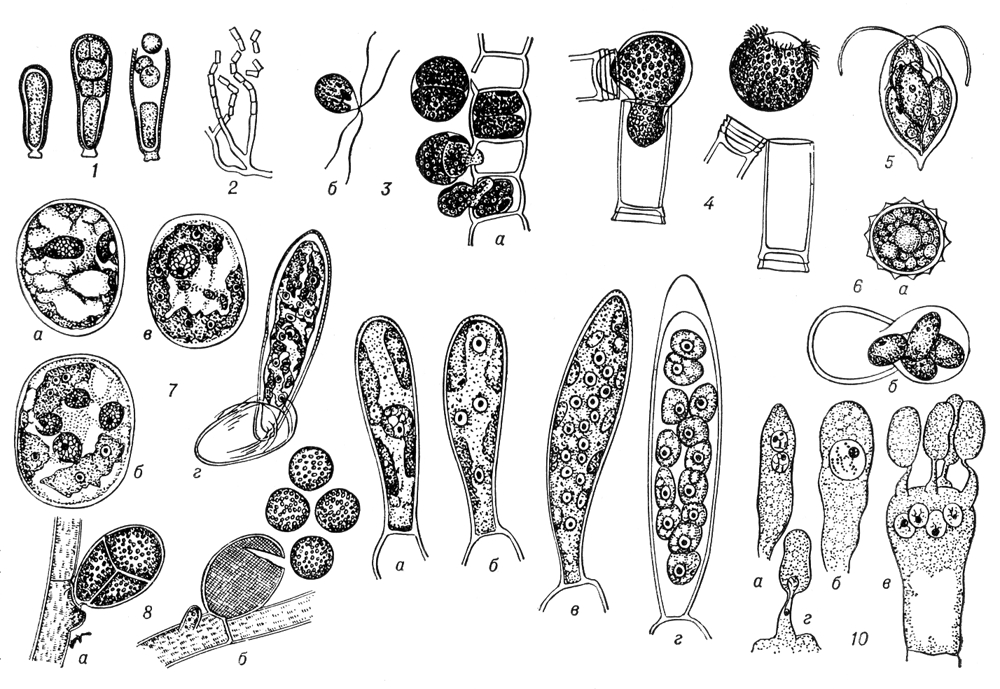
Рис. 1. Спорообразование у низших растений. 1 — образование и выход эндоспор у сине-зеленой водоросли Dermocarpa; 2 — распадение мицелия на членики у актиномицета Nocardia; 3 — улотрикс (Ulothrix): выход спор (а) и спора (б); 4 — эдогониум (Oedogonium): выход зооспоры; 5 — хламидомонада (Chlamydomonas): четыре споры внутри оболочки произведшей их клетки; 6 — хламидомонада (Chlamydomonas): зигота (а) и её прорастание четырьмя спорами (б); 7 — спирогира (Spirogyra): зигота (а) и её прорастание — образование четырёх гаплоидных ядер (б), отмирание трёх ядер (в), одноядерный проросток (г); 8 — каллитамнион (Callithamnion): тетраспорангий (а) и выход тетраспор (б); 9 — ламинариевая водоросль Chorda filum: спорангий с диплоидным ядром (а), четырьмя (б) и шестнадцатью (в) гаплоидными ядрами, с почти созревшими спорами (г); 10 — базидия с дикарионом (а), диплоидным ядром (б) и четырьмя гаплоидными ядрами (в) у базидиальных грибов; г — переход гаплоидного ядра в базидиоспору.