Биологические мембраны, тонкие пограничные структуры молекулярных размеров, расположенные на поверхности клеток и субклеточных частиц, а также канальцев и пузырьков, пронизывающих протоплазму. Толщина Б. м. не превышает 100 . Важнейшая функция Б. м. — регулирование транспорта ионов, сахаров, аминокислот и других продуктов обмена веществ (см. Проницаемость биологических мембран). Первоначально термин «Б. м.» использовали при описании всех видов пограничных структур, встречающихся в живом организме, — покровных тканей, слизистых оболочек желудка и кишечника, стенок кровеносных сосудов и почечных канальцев, миелиновых оболочек нервных волокон, оболочек эритроцитов и др. К середине 20 в. было доказано, что в большинстве пограничных структур эффективную барьерную функцию выполняют не все элементы этих сложных образований, а только мембраны клеток. С помощью электронного микроскопа и рентгеноструктурного анализа удалось показать общность строения поверхностных клеточных мембран эритроцитов, нервных и мышечных клеток, бактерий, плазмалеммы растительных клеток и др. с мембранами субклеточных структур — эндоплазматической сети, митохондрий, клеточных ядер, лизосом, хлоропластов и др. Б.м. занимают огромную площадь (например, в организме человека только поверхностные мембраны имеют площадь, равную десяткам тыс. м2)и играют универсальную регуляторную роль в обмене веществ. Поэтому изучение структуры и функций Б. м. — одна из важнейших задач цитологии и молекулярной биологии. Функции Б. м. многообразны (см. табл.).
Некоторые функции биологических мембран
Функция
Вид мембраны
Активный транспорт веществ
Общая и избирательная диффузия небольших молекул и ионов
Регулирование транспорта ионов и продуктов метаболизма внутри клеток
Все виды мембран
Электроизоляционные свойства
Миелин
Генерация нервного импульса
Мембраны нервных клеток
Преобразование световой энергии в химическую энергию аденозинтрифосфорной кислоты (АТФ)
Мембраны хлоропластов
Преобразование энергии биологического окисления в химическую энергию макроэргических фосфатных связей в молекуле аденозинтрифосфорной кислоты (АТФ)
Мембраны митохондрий
Фагоцитоз, пиноцитоз, антигенные свойства
Мембраны специализированных клеток
Покрывая клетку и отделяя её от окружающей среды, Б. м. обеспечивают морфологическую целостность клеток и субклеточных частиц, их прочность и эластичность. Поддерживая неравномерное распределение ионов калия, натрия, хлора и др. между протоплазмой и окружающей средой, они способствуют появлению разности биоэлектрических потенциалов. Свойства Б. м. в значительной степени определяют генерирование и проведение возбуждения как в нервных и мышечных клетках, так и в местах контакта между ними, т. е. в синаптических окончаниях (см. Синапсы). Б. м. митохондрий служат местом строго упорядоченного расположения ферментов, участвующих в синтезе богатых энергией соединений.
Функциональные свойства Б. м. тесно связаны с их структурной организацией и в значительной степени определяются ею. Ещё в 1902, изучая проницаемость клеточных мембран, немецкий учёный Э. Овертон заметил, что через мембраны легче всего проникают вещества, хорошо растворимые в липидах, и предположил наличие последних в поверхностной клеточной мембране. В 1926 американские биологи Э. Гортер и Ф. Грендел выделили из гемолизированных эритроцитов человека липиды и расположили их в виде мономолекулярного слоя на поверхности воды; общая площадь этого слоя примерно в 2 раза превышала поверхность эритроцитов. Из этого они сделали вывод, что липиды Б. м. расположены в виде бимолекулярного слоя. Поверхностное натяжение клеточной мембраны (0,1 мн/м, или дин/см) меньше натяжения слоя чистого липида (10 мн/м, или дин/см) и близко к поверхностному натяжению белков. Поэтому было предположено, что в Б. м. бимолекулярный липидный слой покрыт с двух сторон слоями белка (структура «сэндвича»). Изучение клеточной поверхности с помощью поляризационного микроскопа позволило предположить, что молекулы липидов расположены перпендикулярно, а молекулы белка — параллельно клеточной поверхности. Методом электропроводности удалось измерить электрическую ёмкость клеточной мембраны, равную 1 мкф/см2, и рассчитать толщину её липидного слоя, которая оказалась равной 55 . На основе всех этих данных английские биологи Л. Даниелли и Г. Даусон в 1935 предложили модель Б. м., в основных чертах удовлетворяющую современным представлениям о структуре Б. м.
Методами рентгеноструктурного анализа, электронной микроскопии, а также оптическими и биохимическими методами показано, что поверхностная клеточная мембрана и мембраны субклеточных частиц — митохондрий, ядер, микросом, лизосом и др. — имеют сходную структуру. Они состоят из бимолекулярного липидного слоя (в основном из фосфолипидов) толщиной 35 и двух нелипидных слоев толщиной 20 каждый (американский исследователь Дж. Робертсон). Внешняя поверхность многих Б. м. покрыта мукополисахаридами. Внутренняя поверхность Б. м. выстлана структурным или ферментным белком (рис. 1, 2). Предполагается, что между молекулами фосфолипидов и белка существует электростатическое притяжение. Мембраны митохондрий несколько отличаются по структуре от поверхностной клеточной мембраны (рис. 3). По-видимому, фосфолипиды и белки в составе внутренней мембраны митохондрий связаны между собой прочным гидрофобным взаимодействием и образуют комплексы («повторяющиеся единицы»), из которых построена вся мембрана.
Значительный прогресс в представлениях о структуре и функции Б. м. достигнут при изучении их моделей — искусственных фосфолипидных мембран, состоящих из бимолекулярного слоя фосфолипидов. Физические свойства такой плёнки близки к свойствам природных Б. м.: толщина её достигает 61 , а электрическая ёмкость 1 мкф/см2. При добавлении в раствор, омывающий искусственную мембрану, небольшого количества белка электрическое сопротивление её резко уменьшается (~ в 1000 раз), приближаясь к электрическому сопротивлению природных Б. м. При определённых условиях в такой «реконструированной» мембране могут возникать электрические колебания, по амплитуде, длительности и условиям возникновения напоминающие электрические колебания в нервном волокне при возбуждении. Добавление в раствор, омывающий эту мембрану, антибиотиков типа валиномицина, грамицидина и др. вызывало появление избирательной проницаемости для ионов калия и натрия. Исследования Б. м. ведутся интенсивно; в ближайшем будущем можно ожидать полной расшифровки их структуры и функции.
Лит.: Руководство по цитологии, т. 1, М.—Л., 1965, гл. 2; Робертис Э. де, Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Робертсон Дж., Мембрана живой клетки, в сборнике: Структура и функция клетки, пер. с англ., М., 1964; Finean J. В., The molecular organization of cell membranes, «Progress in Biophysics and Molecular Biology», 1966, v. 16, p. 143—70.
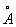 . Важнейшая функция Б. м. — регулирование транспорта ионов, сахаров, аминокислот и других продуктов обмена веществ (см. Проницаемость биологических мембран). Первоначально термин «Б. м.» использовали при описании всех видов пограничных структур, встречающихся в живом организме, — покровных тканей, слизистых оболочек желудка и кишечника, стенок кровеносных сосудов и почечных канальцев, миелиновых оболочек нервных волокон, оболочек эритроцитов и др. К середине 20 в. было доказано, что в большинстве пограничных структур эффективную барьерную функцию выполняют не все элементы этих сложных образований, а только мембраны клеток. С помощью электронного микроскопа и рентгеноструктурного анализа удалось показать общность строения поверхностных клеточных мембран эритроцитов, нервных и мышечных клеток, бактерий, плазмалеммы растительных клеток и др. с мембранами субклеточных структур — эндоплазматической сети, митохондрий, клеточных ядер, лизосом, хлоропластов и др. Б.м. занимают огромную площадь (например, в организме человека только поверхностные мембраны имеют площадь, равную десяткам тыс. м2) и играют универсальную регуляторную роль в обмене веществ. Поэтому изучение структуры и функций Б. м. — одна из важнейших задач цитологии и молекулярной биологии. Функции Б. м. многообразны (см. табл.).
. Важнейшая функция Б. м. — регулирование транспорта ионов, сахаров, аминокислот и других продуктов обмена веществ (см. Проницаемость биологических мембран). Первоначально термин «Б. м.» использовали при описании всех видов пограничных структур, встречающихся в живом организме, — покровных тканей, слизистых оболочек желудка и кишечника, стенок кровеносных сосудов и почечных канальцев, миелиновых оболочек нервных волокон, оболочек эритроцитов и др. К середине 20 в. было доказано, что в большинстве пограничных структур эффективную барьерную функцию выполняют не все элементы этих сложных образований, а только мембраны клеток. С помощью электронного микроскопа и рентгеноструктурного анализа удалось показать общность строения поверхностных клеточных мембран эритроцитов, нервных и мышечных клеток, бактерий, плазмалеммы растительных клеток и др. с мембранами субклеточных структур — эндоплазматической сети, митохондрий, клеточных ядер, лизосом, хлоропластов и др. Б.м. занимают огромную площадь (например, в организме человека только поверхностные мембраны имеют площадь, равную десяткам тыс. м2) и играют универсальную регуляторную роль в обмене веществ. Поэтому изучение структуры и функций Б. м. — одна из важнейших задач цитологии и молекулярной биологии. Функции Б. м. многообразны (см. табл.). . На основе всех этих данных английские биологи Л. Даниелли и Г. Даусон в 1935 предложили модель Б. м., в основных чертах удовлетворяющую современным представлениям о структуре Б. м.
. На основе всех этих данных английские биологи Л. Даниелли и Г. Даусон в 1935 предложили модель Б. м., в основных чертах удовлетворяющую современным представлениям о структуре Б. м. и двух нелипидных слоев толщиной 20
и двух нелипидных слоев толщиной 20 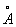 каждый (американский исследователь Дж. Робертсон). Внешняя поверхность многих Б. м. покрыта
каждый (американский исследователь Дж. Робертсон). Внешняя поверхность многих Б. м. покрыта  , а электрическая ёмкость 1 мкф/см2. При добавлении в раствор, омывающий искусственную мембрану, небольшого количества белка электрическое сопротивление её резко уменьшается (~ в 1000 раз), приближаясь к электрическому сопротивлению природных Б. м. При определённых условиях в такой «реконструированной» мембране могут возникать электрические колебания, по амплитуде, длительности и условиям возникновения напоминающие электрические колебания в нервном волокне при возбуждении. Добавление в раствор, омывающий эту мембрану, антибиотиков типа валиномицина, грамицидина и др. вызывало появление избирательной проницаемости для ионов калия и натрия. Исследования Б. м. ведутся интенсивно; в ближайшем будущем можно ожидать полной расшифровки их структуры и функции.
, а электрическая ёмкость 1 мкф/см2. При добавлении в раствор, омывающий искусственную мембрану, небольшого количества белка электрическое сопротивление её резко уменьшается (~ в 1000 раз), приближаясь к электрическому сопротивлению природных Б. м. При определённых условиях в такой «реконструированной» мембране могут возникать электрические колебания, по амплитуде, длительности и условиям возникновения напоминающие электрические колебания в нервном волокне при возбуждении. Добавление в раствор, омывающий эту мембрану, антибиотиков типа валиномицина, грамицидина и др. вызывало появление избирательной проницаемости для ионов калия и натрия. Исследования Б. м. ведутся интенсивно; в ближайшем будущем можно ожидать полной расшифровки их структуры и функции.

