Цитология (от цито... и ...логия), наука о клетке. Ц. изучает клетки многоклеточных животных, растений, ядерно-цитоплазматические комплексы, не расчленённые на клетки (симпласты, синцитии и плазмодии), одноклеточные животные и растительные организмы, а также бактерии. Ц. занимает центральное положение в ряду биологических дисциплин, т.к. клеточные структуры лежат в основе строения, функционирования и индивидуального развития всех живых существ, и, кроме того, она является составной частью гистологии животных, анатомии растений, протистологии и бактериологии.
Развитие цитологиидо начала 20 в. Прогресс Ц. связан с развитием методов исследования клеток. Клеточное строение впервые было обнаружено английским учёным Р. Гуком в ряде растительных тканей в 1665 благодаря использованию микроскопа. До конца 17 в. появились работы микроскопистов М. Мальпиги (Италия), Н. Грю (Великобритания), А. Левенгука (Нидерланды) и др., показавшие, что ткани многих растительных объектов построены из ячеек, или клеток. Левенгук, кроме того, впервые описал эритроциты (1674), одноклеточные организмы (1675, 1681), сперматозоиды позвоночных животных (1677), бактерии (1683). Исследователи 17 в., положившие начало микроскопическому изучению организмов, в клетке видели лишь оболочку, заключающую в себе полость.
В 18 в. конструкция микроскопа была несколько улучшена, главным образом за счёт усовершенствования механических частей и осветительных приспособлений. Техника исследования оставалась примитивной; изучались в основном сухие препараты.
В первые десятилетия 19 в. представления о роли клеток в строении организмов значительно расширились. Благодаря трудам немецких учёных Г. Линка, Я. Мольденхавера, Ф. Мейена, Х. Моля, французских учёных Ш. Мирбеля, П. Тюрпена и др. в ботанике утвердился взгляд на клетки как на структурные единицы. Было обнаружено превращение клеток в проводящие элементы растений. Стали известны низшие одноклеточные растения. На клетки начали смотреть как на индивидуумы, обладающие жизненными свойствами. В 1835 Моль впервые наблюдал деление растительных клеток. Исследования французских учёных А. Мильн-Эдвардса, А. Дютроше, Ф. Распая, чешского учёного Я. Пуркине и др. к середине 30-х гг. дали большой материал по микроскопическим структурам животных тканей. Многие исследователи наблюдали клеточное строение различных органов животных, а некоторые проводили аналогию между элементарными структурами животных и растительных организмов, подготовляя тем самым почву для создания общебиологической клеточной теории. В 1831—33 английский ботаник Р. Броун описал ядро как составную часть клетки. Это открытие привлекло внимание исследователей к содержимому клетки и дало критерий для сопоставления животных и растительных клеток, что и сделал, в частности, Я. Пуркине (1837). Немецкий учёный Т. Шванн, опираясь на теорию развития клеток немецкого ботаника М. Шлейдена, где особое значение придавалось ядру, сформулировал общую клеточную теорию строения и развития животных и растений (1838—39). Вскоре клеточная теория была распространена и на простейших (немецкий учёный К. Зибольд, 1845—48). Создание клеточной теории явилось сильнейшим стимулом к изучению клетки как основы всего живого. Большое значение имело введение в микроскопию иммерсионных объективов (водная иммерсия, 1850, масляная, 1878), конденсора Э. Аббе (1873) и апохроматов (1886). В середине 19 в. начали применяться различные методы фиксации и окраски тканей. Для изготовления срезов были разработаны методы заливки кусочков ткани. Вначале срезы изготовлялись с помощью ручной бритвы, а в 70-х гг. для этого использовались особые приборы — микротомы. В ходе развития клеточной теории постепенно выяснилась ведущая роль содержимого клетки, а не её оболочки. Представление об общности содержимого различных клеток нашло своё выражение в распространении примененного к нему Молем (1844, 1846) термина «протоплазма», введённого Пуркине (1839). Вопреки взглядам Шлейдена и Шванна на возникновение клеток из бесструктурного неклеточного вещества — цитобластемы, с 40-х гг. 19 в. начинает укрепляться убеждение, что умножение числа клеток происходит путём их деления (немецкие учёные К. Негели, Р. Келликер и Р. Ремак). Дальнейшим толчком к развитию Ц. послужило учение немецкого патолога Р. Вирховао «целлюлярной патологии» (1858). Вирхов рассматривал животный организм как совокупность клеток, каждая из которых обладает всеми свойствами жизни; он выдвинул принцип «omnis cellula е cellula» [каждая клетка (происходит только) из клетки]. Выступая против гуморальной теории патологии, которая сводила болезни организмов к порче организменных соков (крови и тканевой жидкости), Вирхов доказывал, что в основе всякого заболевания лежит нарушение жизнедеятельности тех или иных клеток организма. Учение Вирхова заставило патологов заняться изучением клеток. К середине 19 в. «оболочечный» период в изучении клетки заканчивается, и в 1861 работой немецкого учёного М. Шульце утверждается взгляд на клетку как на «комок протоплазмы с лежащим внутри него ядром». В том же году австрийский физиолог Э. Брюкке, считавший клетку элементарным организмом, показал сложность строения протоплазмы. В последней четверти 19 в. был обнаружен ряд постоянных составных частей протоплазмы — органоидов: центросомы (1876, бельгийский учёный Э. ван Бенеден), митохондрии (1897—98, немецкий учёный К. Бенда, у животных; 1904, немецкий учёный Ф. Мевес, у растений), сетчатый аппарат, или комплекс Гольджи (1898, итальянский учёный К. Гольджи). Швейцарский учёный Ф. Мишер (1868) установил в ядрах клеток наличие нуклеиновой кислоты. Открыто кариокинетическое деление клеток (см. Митоз) у растений (1875, Э. Страсбургер), затем у животных (1878, русский учёный П. И. Перемежко; 1882, немецкий учёный В. Флемминг). Создана теория индивидуальности хромосом и установлено правило постоянства их числа (1885, австрийский учёный К. Рабль; 1887, немецкий учёный Т. Бовери). Открыто явление редукции числа хромосом при развитии половых клеток; установлено, что оплодотворение состоит в слиянии ядра яйцевой клетки с ядром сперматозоида (1875, немецкий зоолог О. Гертвиг, у животных; 1880—83, русский ботаник И. Н. Горожанкин, у растений). В 1898 русский цитолог С. Г. Навашин обнаружил у покрытосеменных растений двойное оплодотворение, заключающееся в том, что, помимо соединения ядра спермия с ядром яйцеклетки, ядро второго спермия соединяется с ядром клетки, дающей эндосперм. При размножении растений обнаружено чередование диплоидных (бесполых) и гаплоидных (половых) поколений.
Достигнуты успехи в изучении физиологии клетки. В 1882 И. И. Мечников открыл явление фагоцитоза. Была обнаружена к подробно исследована избирательная проницаемость растительных и животных клеток (голландский учёный Х. Де Фриз, немецкие учёные В. Пфеффер, Э. Овертон); создана мембранная теория проницаемости; разработаны методы прижизненного окрашивания клеток (русский гистолог Н. А. Хржонщевский, 1864; немецкие учёные П. Эрлих, 1885, Пфеффер, 1886). Исследуются реакции клеток на действие раздражителей. Изучение разнообразных клеток высших и низших организмов, несмотря на все их структурные и функциональные различия, укрепило в сознании исследователей мысль о наличии единого принципа в строении протоплазмы. Многие исследователи не были удовлетворены клеточной теорией и признавали наличие в клетках ещё более мелких элементарных жизненных единиц (биобласты Альтмана, пласомы Визнера, протомеры Гейденгайна и т.д.). Спекулятивные представления о субмикроскопических жизненных единицах разделялись и некоторыми цитологами 20 в., однако развитие Ц. заставило большинство учёных оставить эти гипотезы и признать жизнь свойством протоплазмы как сложной гетерогенной системы. Успехи Ц. в конце 19 в. были подытожены в ряде классических сводок, которые способствовали дальнейшему развитию Ц. (Е. В. Wilson, The cell in development and heredity, N. Y., 1928; М. Heidenhain, Plasma und Zeile, 1907; R. Höber, Physikalische Chemie der Zeile und der Gewebe, 1902; М. Verworn, Allgemeine Physiologic, 1895).
Развитие цитологии в 1-й половине 20 в. В первые десятилетия 20 в. стали применять темнопольный конденсор, с помощью которого объекты под микроскопом исследовались при боковом освещении. Темнопольный микроскоп позволил изучать степень дисперсности и гидратации клеточных структур и обнаруживать некоторые структуры субмикроскопических размеров. Поляризационный микроскоп дал возможность определять ориентацию частиц в клеточных структурах. С 1903 развивается микроскопирование в ультрафиолетовых лучах, ставшее в дальнейшем важным методом исследования цитохимии клетки, в частности нуклеиновых кислот. Начинает применяться флюоресцентная микроскопия. В 1941 появляется фазово-контрастный микроскоп, позволяющий различать бесцветные структуры, отличающиеся лишь оптической плотностью или толщиной. Последние два метода оказались особенно ценными при изучении живых клеток. Разрабатываются новые методы цитохимического анализа, среди них — метод выявления дезоксирибо-нуклеиновой кислоты (немецкие учёные Р. Фёльген и Г. Розенбек, 1924). Создаются микроманипуляторы, с помощью которых можно производить над клетками разнообразные операции (инъекции в клетку веществ, извлечение и пересадку ядер, локальное повреждение клеточных структур и т.д.). Большое значение приобрела разработка метода культуры тканей вне организма, начало которому было положено в 1907 американским учёным Р. Гаррисоном. Интересные результаты были получены при сочетании этого метода с замедленной микрокиносъёмкой, что дало возможность видеть на экране медленные изменения в клетках, протекающие незаметно для глаза, ускоренными в десятки и сотни раз. В первые три десятилетия 20 в. усилия учёных направлены были на выяснение функциональной роли клеточных структур, открытых в последней четверти 19 в., в частности было установлено участие комплекса Гольджи в выработке секретов и др. веществ в гранулярной форме (советский учёный Д. Н. Насонов, 1923). Описаны частные органоиды специализированных клеток, опорные элементы в ряде клеток (Н. К. Кольцов, 1903—1911), исследованы структурные изменения при различной клеточной деятельности (секреция, сократительная функция, деление клеток, морфогенез структур и т.д.). В растительных клетках прослежено развитие вакуолярной системы, образование крахмала в пластидах (французский учёный А. Гийермон, 1911). Установлена видовая специфичность числа и формы хромосом, что в дальнейшем было использовано для систематики растений и животных, а также для выяснения филогенетического родства в пределах более низких таксономических единиц (кариосистематика). Обнаружено, что в тканях имеются разные классы клеток, отличающихся кратным отношением размеров ядер (немецкий учёный В. Якоби, 1925). Кратное увеличение размера ядер сопровождается соответствующим увеличением (путём эндомитоза) числа хромосом (австрийский учёный Л. Гейтлер, 1941). Исследования действия агентов, нарушающих механизм деления и хромосомный аппарат клеток (проникающее излучение, колхицин, ацетонафтен, трипофлавин и др.), привели к разработке методов искусственного получения полиплоидных форм (см. Полиплоидия), что дало возможность вывести ряд ценных сортов культурных растений. С помощью реакции Фёльгена положительно решился спорный вопрос о наличии гомолога ядра, содержащего дезоксирибонуклеиновую кислоту у бактерий (советский учёный М. А. Пешков, 1939—1943, французский учёный В. Делапорт, 1939, английский учёный С. Робиноу, 1942) и сине-зелёных водорослей (советские учёные Ю. И. Полянский и Ю. К. Петрушевский, 1929). Наряду с мембранной теорией проницаемости, выдвигается фазовая теория, придающая большое значение в распределении веществ между клеткой и средой, растворению их и связыванию в протоплазме (советские учёные Д. Н. Насонов, В. Я. Александров, А. С. Трошин). Изучение реакции протоплазмы клеток на воздействие разнообразных физических и химических агентов привело к обнаружению явлений паранекроза и к разработке денатурационной теории повреждения и возбуждения (Д. Н. Насонов и В. Я. Александров, 1940), согласно которой в этих процессах ведущее значение имеют обратимые изменения в структуре белков протоплазмы. С помощью вновь разработанных цитохимических реакций на гистологических препаратах была установлена локализация в клетке ряда ферментов. Начиная с 1934 благодаря работам американских учёных Р. Уэнсли и М. Герр, использовавшим метод гомогенизации (размельчения) клеток и фракционного центрифугирования, началось извлечение из клеток отдельных компонентов — ядер, хлоропластов, митохондрий, микросом и изучение их химического и ферментативного состава. Однако существенные успехи в расшифровке функций клеточных структур достигнуты лишь в современный период развития Ц. — после 50-х гг.
Огромное влияние на развитие Ц. в 20 в. оказало переоткрытие в 1900 Менделя законов. Изучение процессов, протекающих в ядрах половых и соматически клеток, дало возможность объяснить факты, установленные при изучении наследственной передачи признаков, и построить хромосомную теорию наследственности. Изучение цитологических основ наследственности обособилось в отдельную отрасль Ц. — цитогенетику.
Развитие современной цитологии. С 50-х гг. 20 в. Ц. вступила в современный этап своего развития. Разработка новых методов исследования и успехи смежных дисциплин дали толчок бурному развитию Ц. и привели к стиранию чётких границ между Ц., биохимией, биофизикой и молекулярной биологией. Использование электронного микроскопа (его разрешающая способность достигает 2—4 , предел разрешения светового микроскопа около 2000 ) привело к созданию субмикроскопической морфологии клетки и приблизило визуальное изучение клеточных структур к макромолекулярному уровню. Были обнаружены неизвестные до этого детали строения ранее открытых клеточных органоидов и ядерных структур; открыты новые ультрамикроскопические компоненты клетки: плазматическая, или клеточная, мембрана, отграничивающая клетку от окружающей среды, эндоплазматический ретикулум (сеть), рибосомы (осуществляющие синтез белка), лизосомы (содержащие гидролитические ферменты), пероксисомы (содержащие ферменты каталазу и уриказу), микротрубочки и микрофиламенты (играющие роль в поддержании формы и в обеспечении подвижности клеточных структур); в растительных клетках обнаружены диктиосомы — элементы комплекса Гольджи. Наряду с общеклеточными структурами выявляются ультрамикроскопические элементы и особенности, присущие специализированным клеткам. С помощью электронной микроскопии показано особое значение мембранных структур в построении различных компонентов клетки. Субмикроскопические исследования дали возможность все известные клетки (и соответственно все организмы) разделить на 2 группы: эукариоты (тканевые клетки всех многоклеточных организмов и одноклеточные животные и растения) и прокариоты (бактерии, сине-зелёные водоросли, актиномицеты и риккетсии). Прокариоты — примитивные клетки — отличаются от эукариотов отсутствием типичного ядра, лишены ядрышка, ядерной оболочки, типичных хромосом, митохондрий, комплекса Гольджи.
Усовершенствование методов изоляции клеточных компонентов, использование методов аналитической и динамической биохимии применительно к задачам Ц. (меченные радиоактивными изотопами предшественники, авторадиография, количественная цитохимия с использованием цитофотометрии, разработка цитохимических методик для электронной микроскопии, применение антител, меченных флуорохромами, для обнаружения под флуоресцентным микроскопом локализации индивидуальных белков; метод гибридизации на срезах и мазках радиоактивных ДНК и РНК для идентификации нуклеиновых кислот клетки и т.д.) привело к уточнению химической топографии клеток и расшифровке функционального значения и биохимической роли многих составных частей клетки. Это потребовало широкого объединения работ в области Ц. с работами по биохимии, биофизике и молекулярной биологии. Для изучения генетических функций клеток большое значение имело открытие содержания ДНК не только в ядре, но и в цитоплазматических элементах клетки — митохондриях, хлоропластах, а по некоторым данным, и в базальных тельцах. Для оценки роли ядерного и цитоплазматического генного аппарата в определении наследственных свойств клетки используется пересадка ядер и митохондрий. Гибридизация соматических клеток становится перспективным методом изучения генного состава отдельных хромосом (см. Соматических клеток генетика). Установлено, что проникновение веществ в клетку и в клеточные органоиды осуществляется с помощью особых транспортных систем, обеспечивающих проницаемость биологических мембран. Электронно-микроскопические, биохимические и генетические исследования увеличили число сторонников гипотезы симбиотического (см. Симбиогенез) происхождения митохондрий и хлоропластов, выдвинутой в конце 19 в.
Основные задачи современной Ц. — дальнейшее изучение микроскопических и субмикроскопических структур и химической организации клеток; функций клеточных структур и их взаимодействий; способов проникновения веществ в клетку, выделения их из клетки и роли мембран в этих процессах; реакций клеток на нервные и гуморальные стимулы макроорганизма и на стимулы окружающей среды; восприятия и проведения возбуждения; взаимодействия между клетками; реакций клеток на повреждающие воздействия; репараций повреждения и адаптации к факторам среды и повреждающим агентам; репродукции клеток и клеточных структур; преобразований клеток в процессе морфофизиологической специализации (дифференцировки); ядерного и цитоплазматического генетического аппарата клетки, его изменений при наследственных заболеваниях; взаимоотношений клеток с вирусами; превращений нормальных клеток в раковые (малигнизация); процессов поведения клеток; происхождения и эволюции клеточной системы. Наряду с решением теоретических вопросов Ц. участвует в разрешении ряда важнейших биологических, медицинских и с.-х. проблем. В зависимости от объектов и методов исследования развивается ряд разделов Ц.: цитогенетика, кариосистематика, цитоэкология, радиационная Ц., онкологическая Ц., иммуноцитология и т.д.
В СССР имеются специальные цитологические исследовательские учреждения: институт цитологии АН СССР, институт цитологии и генетики Сибирского отделения АН СССР, институт генетики и цитологии АН БССР. Во многих др. биологических, медицинских и с.-х. научных учреждениях имеются специальные цитологические лаборатории. Работы по Ц. координируются в СССР Научным советом по проблемам Ц. при АН СССР. Издаются журналы «Цитология» (АН СССР), «Цитология и генетика» (АН УССР). Цитологические работы публикуются в журналах по смежным дисциплинам. В мире издаётся более 40 цитологических журналов. Периодически выходят книги многотомных интернациональных изданий: протоплазматология («Protoplasmatologia») (Вена) и международное обозрение по Ц. («International Review of Cytology») (Нью-Йорк). Имеется Международное общество биологии клетки (International Society of Cell Biology), регулярно созывающее цитологические конгрессы. Международная организация по исследованию клетки (International Cell Research Organization) и Европейская организация по биологии клетки (European Cell Biology Organization) создают рабочие группы по отдельным проблемам Ц., организуют курсы по узловым вопросам Ц. и для изучения методик, обеспечивают обмен информацией. В университетах СССР на биологических и биолого-почвенных факультетах преподаётся курс общей Ц. Во многих университетах проводятся специализированные курсы по разным проблемам Ц. В виде раздела Ц. входит также в состав курсов гистологии животных, анатомии растений, эмбриологии, протистологии, бактериологии, физиологии, патологической анатомии, которые читаются в с.-х., педагогических и медицинских учебных заведениях. См. также ст. Клетка и лит. при ней.
Лит.: Кацнельсон З. С., Клеточная теория в ее историческом развитии, Л., 1963; Руководство по цитологии, т. 1—2, М. — Л., 1965—66; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1973; Brown W. V., Bertke E. M., Textbook of cytology, Saint Louis, 1969; Hirsch G. С., Ruska H., Sitte P., Grundlagen der Cytologie, Jena, 1973.
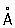 , предел разрешения светового микроскопа около 2000
, предел разрешения светового микроскопа около 2000 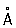 ) привело к созданию субмикроскопической морфологии клетки и приблизило визуальное изучение клеточных структур к макромолекулярному уровню. Были обнаружены неизвестные до этого детали строения ранее открытых клеточных органоидов и ядерных структур; открыты новые ультрамикроскопические компоненты клетки: плазматическая, или клеточная, мембрана, отграничивающая клетку от окружающей среды, эндоплазматический ретикулум (сеть), рибосомы (осуществляющие синтез белка), лизосомы (содержащие гидролитические ферменты), пероксисомы (содержащие ферменты каталазу и уриказу), микротрубочки и микрофиламенты (играющие роль в поддержании формы и в обеспечении подвижности клеточных структур); в растительных клетках обнаружены диктиосомы — элементы комплекса Гольджи. Наряду с общеклеточными структурами выявляются ультрамикроскопические элементы и особенности, присущие специализированным клеткам. С помощью электронной микроскопии показано особое значение мембранных структур в построении различных компонентов клетки. Субмикроскопические исследования дали возможность все известные клетки (и соответственно все организмы) разделить на 2 группы: эукариоты (тканевые клетки всех многоклеточных организмов и одноклеточные животные и растения) и прокариоты (бактерии, сине-зелёные водоросли, актиномицеты и риккетсии). Прокариоты — примитивные клетки — отличаются от эукариотов отсутствием типичного ядра, лишены ядрышка, ядерной оболочки, типичных хромосом, митохондрий, комплекса Гольджи.
) привело к созданию субмикроскопической морфологии клетки и приблизило визуальное изучение клеточных структур к макромолекулярному уровню. Были обнаружены неизвестные до этого детали строения ранее открытых клеточных органоидов и ядерных структур; открыты новые ультрамикроскопические компоненты клетки: плазматическая, или клеточная, мембрана, отграничивающая клетку от окружающей среды, эндоплазматический ретикулум (сеть), рибосомы (осуществляющие синтез белка), лизосомы (содержащие гидролитические ферменты), пероксисомы (содержащие ферменты каталазу и уриказу), микротрубочки и микрофиламенты (играющие роль в поддержании формы и в обеспечении подвижности клеточных структур); в растительных клетках обнаружены диктиосомы — элементы комплекса Гольджи. Наряду с общеклеточными структурами выявляются ультрамикроскопические элементы и особенности, присущие специализированным клеткам. С помощью электронной микроскопии показано особое значение мембранных структур в построении различных компонентов клетки. Субмикроскопические исследования дали возможность все известные клетки (и соответственно все организмы) разделить на 2 группы: эукариоты (тканевые клетки всех многоклеточных организмов и одноклеточные животные и растения) и прокариоты (бактерии, сине-зелёные водоросли, актиномицеты и риккетсии). Прокариоты — примитивные клетки — отличаются от эукариотов отсутствием типичного ядра, лишены ядрышка, ядерной оболочки, типичных хромосом, митохондрий, комплекса Гольджи.