листовых органов. Листья таких тенелюбов, как кислица (Oxalis Acetosella), имеют мягкость, составленную исключительно из клеток губчатой паренхимы (редко расположенных), и наоборот, у растений весьма светолюбивых она составлена почти исключительно из клеток столбчатой, или палисадной, ткани (плотно расположенных). Те же исследования показали, что листья некоторых растений, в зависимости от того, родились ли они на свету или в тени, имеют паренхиму, то составленную преимущественно из губчатой ткани, то наоборот, – из столбчатой. Как известно, ткань губчатая приспособлена для работы при ослабленном свете, палисадная же – для более сильного освещения. На рис. 54 и 55, заимствованных из работ Шталя, можно видеть, как варьирует лист бука в зависимости от того, находится ли он на периферии кроны или внутри ее. На рис. 56 и 57 можно видеть большую пластичность хвои другой древесной породы, именно ели. Вот этими-то наблюдениями над степенью пластичности внутреннего и внешнего строения листовых органов и воспользовался лесовод И. И. Сурож, применив эти начала к исследованию большого числа древесных пород. Измеряя на поперечном разрезе листа длину губчатой и палисадной- ткани у разных пород, названный исследователь обнаружил весьма различную пластичность их: у одних – в тени более сильно, у других – более слабо развита палисадная ткань, и чем она в тени менее развита, тем большей способностью приспособляться отличается данная
Порода. Не входя в рассмотрение методики, мы ограничимся только приведением той шкалы теневыносливости, в какую И. И. Сурож расположил древесные породы, причем ограничимся только важнейшими из исследованных им пород. По строению ассимилятивной ткани, по И. И. Сурожу, наши ло-
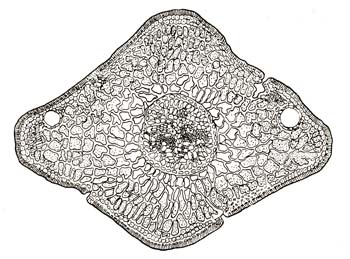
Рис. 56. Разрез через хвою ели, выросшей на свету
роды могут быть расположены, начиная с самых теневыносливых, в следующий ряд:
тисе,
пихта,
липа,
клен,
черная ольха,
пушистая береза,
вяз,
белая ольха,
дуб летний,
ясень,
береза бородавчатая,
осина,
кедр сибирский,
сосна обыкновенная,
лиственница,
сосна горная.
Лесовод и ботаник В. Н. Любименко видит причину различной теневыносливости в различной чувствительности хлорофиллоносного аппарата к свету. Оперируя с некоторыми древесными породами, он показал, что ассимилятивная деятельность листовых органов начинается у разных пород при различной интенсивности света, и что именно теневыносливые породы требуют для этого значительно меньшей силы света, чем породы светолюбивые. В особый ящик, имевший боковое отверстие с матовым стеклом, которое можно было притом увеличивать и уменьшать, помещались две пробирки, одна из которых – зачерненная лаком; в эти пробирки вкладывались отрезки листьев следующих растений: обыкновенной сосны, пихты, липы и березы. – в одном опыте, тисса, лиственницы, бука и белой акации – в другом. Пользуясь искусственным освещением, регулируя его силу различной величиной отверстия и производя анализ газов в пробирках после опыта, В. Н. Любименко мог показать, что сосна, например, начинала усвоение углекислоты только при величине отверстия в 49 см2, береза – при 64, тогда как пихта и липа – при 9 см2, бук – при 4, акация – при 100 см2. Иначе говоря, хлорофилломонный аппарат пихты и липы приблизительно в 5 раз
чувствительнее такового же сосны и березы; бук требует для начала ассимиляции напряженности света в 25 раз, а тисе в 11 раз более слабой, чем лиственница и белая акация. Спектроскопические исследования показали тому же автору, что растения теневыносливые характеризуются большей концентрацией хлорофилла, чем светолюбивые. У первых, кроме того, хлоропласты крупнее, чем у вторых. На рис. 58, 59, 60, 61, сделанных при одинаковом увеличении, например, видно, что у тисе а хлоропласты гораздо крупнее, чем у лиственницы, а у бука значительно крупнее, чем у белой акации.
Третий метод, впервые примененный проф. М. К. Турским, основан на другом явлении, известном из физиологии растений, именно этиолировании. В работе его ученика В. Никольского было прослежено влияние отенения на рост однолетней сосны и ели. Эти растения были высеяны на грядах, из коих одна была оставлена свободной, другие же были затенены

Рис. 57. Разрез через хвою ели, выросшей в тени.
щитами, у которых промежутки между дранками были разной величины, именно на одной гряде промежутки между дранками равнялись ширине их, или освещение считали равным 1/2, на другой расстояния равнялись ширины дранок, или освещение рав-

Рис. 58. Хлоропласта тисса (в клетках листа)

Рис. 60. Хлоропласта бука

Рис. 59. Хлоропласта лиственницы

Рис. 61. Хлоропласта белой акации
нялось 1/3. По окончании опыта были измерены длина и вес сухого вещества надземных и подземных частей. Оказалось что как длина главного корня, так и число, и общая длина боковых, с увеличением отенения уменьшаются у сосны в 7 1/2 раз, у ели – почти в 2 раза; длина же всей надземной части как и надо было ожидать по законам физиологии растений, как у сосны, так и у ели с усилением отенения увеличивается. Вес сухого вещества сеянцев на полном свету, как опять-таки надо было ожидать, оказался больше веса сухого вещества сеянцев, выросших при сильном отенении: у сосны почти в 5 раз, у ели почти в 3 раза.
Этот опыт, опубликованный в 1881 г. и произведенный, к сожалению, только над двумя породами, недавно повторен известным австрийским лесоводом Цизляром в несколько иной форме и над большим числом пород. На грядах почти тем же способом было достигнуто семь степеней отенения, но эффект отенения измерялся не весом сухого вещества, а объемом растения. Если объем 100 сеянцев, выросших при полном освещении, выразить величиной 100, то объемы, полученные при меньшем освещении, выразятся следующими величинами (табл. 9):
Таблица 9
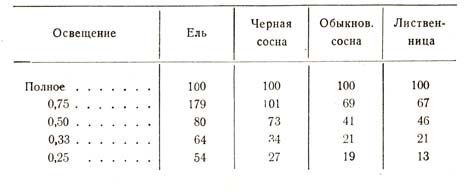
Мы видим, что при наибольшем затенении лиственница пострадала более всего, дав только 13% объема от объема ее при полном освещении. Затем следует обыкновенная сосна, далее – черная сосна и, наконец, ель, которую поэтому и следует признать самой теневыносливой из названных пород, а лиственницу – самой светолюбивой.
Измерение длины годичного побега точно так же показало, что отенение отразилось более всего на лиственнице, сосне обыкновенной и, наконец, на ели: у первой длина выразилась в цифре 176, у второй – 155 и у третьей – 133, вместо 100 при нормальных условиях.
Наш известный лесовод и ботаник Я. С. Медведев, желая избавиться от субъективности в оценке светолюбия породы, предложил свой способ измерения и вычисления относительной высоты деревьев, допускающий объективную оценку. Сущность этого метода, помещенного в «Лесном журнале» за 1884 г. в статье, озаглавленной «К учению о влиянии света на развитие древесных стволов», заключается в следующем: дерево, свободно растущее, растет преимущественно в толщину; дерево среди насаждения, напротив, растет преимущественно в высоту; таким образом, между ростом дерева в высоту и в толщину существует известное соотношение, зависящее от степени освещения; несомненно, что отношение высоты дерева к его диаметру будет наименьшим у дерев, выросших на свободе, и наибольшим у дерев, растущих в густых насаждениях, притом в состоянии угнетения; среднее же место будут занимать господствующие деревья насаждения. Положение это, вытекающее уже a priori из закона распределения прирастающей массы по дереву, подтверждено Медведевым, а затем Марченко на основании данных многочисленных пробных площадей. Назвав это отношение высоты дерева к его диаметру, выраженным притом в одной и той же мере, относительной высотой, Медведев нашел следующие предельные величины (табл. 10):
Таблица 10

Между этими пределами заключаются относительные высоты всех промежуточных степеней освещения. Такой характер изменений в росте древесных стволов в зависимости от количества падающего на них света, по мнению Медведева, дает возможность пользоваться относительной высотой как самым чувствительным измерителем степени освещения деревьев. Чем древесная порода более теневынослива, тем полнее угнетение, за которым наступает смерть дерева; она наступает при большем недостатке света и, стало быть, при более высокой относительной высоте. Последняя дает возможность определить с точностью сравнительную способность древесных пород переносить недостаток света, т. е. теневыносливость. Лучший прием для этого заключался бы, по мнению Медведева, в вычислении относительной высоты дерева, выросшего вне взаимного влияния, но под совершенно одинаковым отенением какого-нибудь насаждения. Не располагая таким материалом, автор этого метода воспользовался многочисленными (результатами своей таксационной практики, обширным материалом измерения пробных площадей в лесах Закавказья. Не останавливаясь на подробностях метода и на тех условиях, соблюдение которых гарантировало автору правильность выводов, приведу лишь некоторые фактические данные и заключительные выводы.
Наибольшее число измерений сделано для ели (Picea oricn-talisj и сосны (Pinus silvestris L.), собранных в виде таблицы (табл. ,11).
Таблица 11
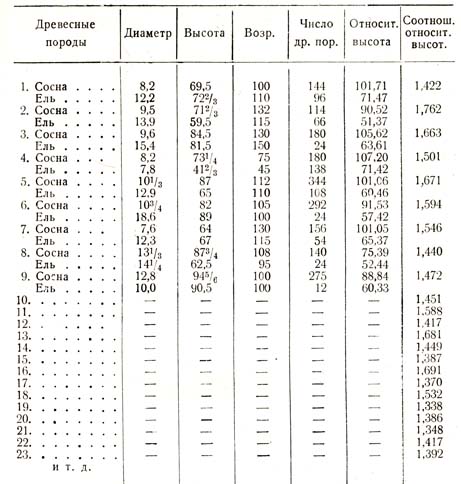
В этой таблице показано, кроме найденных относительных высот, и взаимные отношения их, крайние пределы которых колеблются между 1,762 и 1,338; средняя величина определяется для ели цифрой 1,500, для сосны обратной дробью – 0,666, согласно чему теневыносливость ели в полтора раза более, или, наоборот, сосна способна выносить только 2/3 того отенения, которое выносит ель. Теневыносливость березы определена по отношению к сосне на основании данных четырех пробных площадей. Беря среднее из полученных соотношений относительных высот березы к сосне, получаем:
(1,143-4-1,188+ 1.5234 1,357)/4=1,303
Световые отношения бука определены по сравнению с елью на основании восьми пробных площадей, причем средняя соотношений между относительными высотами этих пород найдена равной 1,029. Таким путем были определены световые отношения других пород, причем были получены следующие величины для выражения световых отношений: ели к сосне – как 1,500, сосны к березе – 1,333, бука к ели – 1,029, граба к буку – 0,918, тисса к буку – 2,816, пихты к ели – 1,125, липы к буку – 0,849, дуба к грабу – 0,855, осины к сосне – 1,199 и ясеня к березе – 1,400.
Принявши затем березу, как самую светолюбивую породу, за единицу, Медведев получил следующие величины для выражения относительной теневыносливости древесных пород, приведенных к единице, именно:
Береза........ 1,000
Сосна........ 1,333
Ясень........ 1,400
Осина........ 1,598
Дуб......... 1,645
Липа........ 1,747
Граб ....... 1,889
Ель......... 2,000
Бук......... 2,058
Пихта ...... 2,250 (Кавказ)
Тисе......... 5,795
Исследования Марченко, имевшие предметом относительную высоту деревьев, показали, что, тогда как угнетенные деревья прирастают в толщину в 2,3 – 3,3 раза медленнее, чем господствующие экземпляры, первые отстают в высоту от последних лишь в 1,3 – 1,5 раза. Исследования Р. Гартига показали, что прирастающая масса распределяется по дереву различно, в зависимости от степени сомкнутости насаждения и принадлежности к тому или иному классу; тогда как у господствующих деревьев прирост откладывается в нижней части ствола, у угнетенных, наоборот, в верхних частях; констатированы даже случаи совершенного выпадения годичных колец на высоте груди у угнетенных деревьев. Неизбежным следствием такого развития деревьев господствующих и угнетенных должна являться более значительная относительная высота последних, что на самом деле и наблюдается. Подтверждая своими фактическими данными положение Медведева об изменяемости относительной высоты в зависимости от принадлежности дерева к тому или иному классу, Марченко совершенно справедливо ставит вопрос о том, можно ли это изменение поставить в зависимость лишь от условий освещения. Для положительного ответа на этот вопрос надо доказать, что распределение прирастающей массы по дереву зависит исключительно или главным образом от условий пользования светом, т. е. степени освещения, выше было упомянуто, что борьба за существование между деревьями какого-либо насаждения, обусловливающая дифференциацию их на господствующие и угнетенные классы, едва ли может рассматриваться как исключительно борьба их за свет.
Сказанным нам приходится ограничить тот материал, который имеется в распоряжении лесоводов. Так или иначе, но мы располагаем и целым рядом методов, и целым рядом шкал теневыносливости наших важнейших древесных пород, которыми и можно пользоваться при наблюдениях над жизнью леса.
Степень теневыносливости какой-нибудь породы, будучи ее видовой особенностью, не является, однако, величиной неизменной. Давно лесоводством установлено, что потребность в свете у одной и той же породы изменяется в зависимости от возраста, почвы и климата. Существует положение, что с возрастом каждая порода становится светолюбивее [1 Академиком Т. Д. Лысенко разработано учение о стадийном развитии растений, согласно которого растения переживают качественные стадии своего развития, характеризующиеся разными требованиями к условиям среды. Данное учение открыло новые широкие возможности для изучения природы растений и их переделки. Ред,]. В таком виде это положение справедливо оспаривается Я. С. Медведевым. Правильнее сказать, что с увеличением возраста увеличивается потребность в большем просторе. Другое положение, – что с улучшением почвенных условий увеличивается теневыносливость породы, – тоже еще требует доказательств.
Третье положение – лесоводственное – гласит, что теневыносливость породы увеличивается с улучшением климатических условий. Это старинное положение формулировано лесоводом Майром таким образом, что каждая порода, обладая в оптимуме своего роста определенной теневыносливостью, увеличивает таковую по направлению к более теплому району своего распространения и, наоборот, становится более светолюбивой в более холодной части своей зоны. Наблюдения его показали, например, что ясень, сосна, лиственница, являясь в своем климатическом оптимуме породами светолюбивыми, в более теплом климате становятся полутеневыносливыми и, наоборот, такие теневыносливые породы, как пихта и бук, в более холодном климате становятся и более светолюбивыми. Эта очень вероятная закономерность более точных объективных подтверждений пока еще не имеет. Если это положение верно, то мы должны принять его во внимание и при постановке тех опытов, которыми имеется в виду выяснить степень теневыносливости разных пород. На самом деле, если мы, например, совершенно точно определим степень этиолирования разных пород при одной и той же степени отенения в ленинградском климате, то, возможно, что такой же опыт, проведенный в Харьковском округе, может дать другие результаты; так, ель, например, в условиях ленинградского климата может быть ближе к своему оптимуму, чем в Харьковском окр., где она совсем не встречается; наоборот, дуб в Харьковском окр., хотя еще далеко расположен о г своего оптимума, но все же ближе к нему, чем дуб Ленинградской области.
Чтобы покончить с отношением пород к свету, необходимо указать, хотя и совершенно кратко, на лесообразовательную роль потребности древесных пород в свете.
Не может быть никакого сомнения, что степень теневыносливости должна влиять на ход борьбы за существование между породами. Сейчас мы не имеем данных, чтобы определить степень этого влияния или чтобы утверждать, как это иногда делают, что теневыносливость определяет даже исход борьбы за существование. Последнее ошибочно: как бы ни была велика теневыносливость ели по сравнению с сосною, ель ничего от того не выигрывает, если почва не будет соответствовать ели; ель будет влачить жалкое существование, будет низкорослой, хронически больным, хронически голодающим организмом. На подобных, не отвечающих ели по своей бедности или сухости, почвах последняя не бывает долговечным растением: она почти не цветет и не плодоносит. Поселившись в подобных условиях под пологом соснового леса, ель будет мешать сосне, в особенности возобновлению этой породы, но все же не вытеснит отсюда сосны. Однако с улучшением почвенных условий все большую роль в борьбе этих пород будет играть теневыносливость ели и светолюбив сосны. Пока мы можем сказать только, что при прочих равных условиях та порода будет иметь больше шансов на победу в борьбе за существование, которая отличается большей теневыносливостью. Но и это положение не может иметь абсолютного значения, – оно весьма относительное, так как необходимо принять во внимание ту степень соответствия климатическим и почвенным условиям, какая наблюдается для разных пород, образующих в том или другом месте лесное сообщество. Будь иначе, не могли бы удержаться в наших лесах светолюбивые элементы. Они должны были бы за четвертый период исчезнуть с лица земли, не выдержав борьбы с породами теневыносливыми. Мы видим, однако, другое: в наших дубравах сохранилась такая светолюбивая порода, как дуб, в сочетании с такими теневыносливыми, как клены, граб, бук, липа; среди наших ельников сохранились такие светолюбивые элементы, как осина и береза. Все это приводится не для умаления значения теневыносливости, но для указания на то, что нельзя и преувеличивать роль этого фактора. Не могу не обратить еще внимания на то, что в каждой флористической области и в каждой области роста существуют свои тенелюбы и свои светолюбы, притом порознь на разных участках, а часто в одном и том же сообществе, притом довольно устойчиво, как бы в стадии подвижного равновесия. Есть, стало быть, какие-то еще условия и какие-то другие факторы, которые содействую светолюбивым элементам или препятствуют теневыносливым породам праздновать всеобщую победу.
Так или иначе, но вне всякого сомнения, что теневыносливость пород имеет то или иное, в разных случаях разное, но бесспорное влияние на ход борьбы за существование между породами. Это справедливо не только по отношению к разным породам, борющимся между собою в одном и том же сообществе, но и по отношению к отдельным членам одного и того же чистого насаждения, так как в силу закона индивидуальности, всюду наблюдаемого в живом мире, одни экземпляры одного и того же 1вида могут быть светолюбивее, другие теневыносливее своих собратьев. Эта видовая, передаваемая по наследству, особенность, наверно, варьирует в известных пределах между отдельными особями одного и того же вида, не говоря уже о расах и разновидностях.
Мы увидим впоследствии, что любая биологическая особенность какой-либо породы не стоит особняком, а находится в тесной и закономерной связи с другими биологическими особенностями, так что сумма их, создающая определенную лесоводственную физиономию той или иной породы, есть как бы биологический аккорд, есть гармоническое сочетание различных особенностей, выработавшихся под влиянием внешних условий борьбы за существование и естественного отбора; сумма биологических свойств породы не есть сумбурный ряд случайных привычек и свойств. Мы увидим, что с светолюбием связана быстрота роста, определенный тип плодоношения, малая чувствительность к заморозкам и т. д. Увидим впоследствии, что наши важнейшие древесные породы можно будет расчленить на несколько типов лесообразователей, например на породы-пионеры и так называемые основные породы, причем светолюбие играет здесь большую роль. Пионеры светолюбивы, быстрорастущи, нечувствительны к заморозкам, обильно и часто плодоносящи с легкими семенами, с определенными чертами вегетативного размножения и т. д. (например, осина, береза, белая ольха и др.). Основные же породы теневыносливы, с медленным ростом в молодости, чувствительны к заморозкам, с более тяжелыми семенами, с более редким плодоношением (например, бук, ель, пихта, липа и т. д.). Есть и двуликие Янусы, как, например, обыкновенная сосна, которая обладает и свойствами пионера, и свойствами основной лесной породы, являясь в одних случаях пионером леса, в других – основной лесной породой.
Ясное дело, что там, где территорию надо еще завоевать, где много инсоляции, но вместе и излучения тепла, где готова разрастись густая дернина светолюбивых трав и злаков, где быстро надо затянуть большое пространство лесом, – там все преимущества будут иметь пионеры, как они своим быстрым ростом скоро справятся с травою, так как они своим обильным плодоношением и легкими семенами скоро покроют всю площадь.
Одним словом, и у светолюбивых элементов есть свои преимущества по сравнению с теневыносливыми три определенных условиях внешней среды.
Какую же еще роль играют светолюбив и теневыносливость в лесообразовании и в жизни леса? Светолюбивые породы образуют сообщества, в которых процесс борьбы за существование, как показывают все опытные таблицы в лесоводстве, совершается гораздо быстрее, чем в насаждениях из пород теневыносливых. Я не буду приводить доказательств этому, – их читатель найдет в первой части моей книги. В насаждениях чистых раньше выделяются угнетенные классы, быстрее проходят они все ступени от первоначального небольшого заглушения до окончательной смерти. Благодаря более быстрому процессу изреживания, число стволов на гектаре в насаждениях из светолюбивых пород всегда меньше, чем в том же возрасте в сообществе пород теневыносливых. Благодаря более раннему и более сильному процессу естественного изреживания в насаждениях пород светолюбивых, последние в меньшей степени преобразующе влияют на занятую среду, чем сообщество теневыносливых пород. В первом случае сквозь полог насаждения больше проникает осадков и больше проходит света и тепла. Благодаря менее густой кроне светолюбивых пород, ежегодный отпад хвои, листьев и мелких веток не достигает тех размеров, в каких он бывает в ельниках и тому подобных насаждениях с мощным мертвым покровом. Травяной покров отличается более светолюбивым характером, влажность и температура воздуха под пологом не так резку отличаются от соответствующих элементов свободного пространства, как при сравнении атмосферы под пологом леса из теневой породы – со свободной поляной.
Породы светолюбивые в силу этого своего свойства, если почвенные условия не препятствуют, должны образовывать смешанные и сложные насаждения, так как, быстро и сильно изреживаясь, они легко дают приют другим элементам – или столь же светолюбивым, или более теневыносливым. Не будь разной степени теневыносливости, не было бы на свете и сложных насаждений, где породы распределены по ярусам в зависимости от степени теневыносливости.
Итак, мы видели, что изучаемые нами свойства пород должны иметь большое значение в их совместной, или общественной жизни. И борьба за существование – ее интенсивность и ход, и самоизреживание насаждений, и появление самосева и подроста, или, иначе, самовозобновление леса, и способность леса преобразовывать внешнюю среду, в частности количество, характер и состав живого покрова в лесу, и способность породы образовывать чистые или смешанные, простые или сложные сообщества, – все это и многое другое в жизни и формах леса находится наряду с другими факторами в тесной связи с теневыносливостью пород.
Можно было бы еще остановиться на роли светолюбия пород при метаморфозах леса, или так называемой смене пород, но этому чрезвычайно сложному и важному явлению будет посвящен особый отдел в книге.
ОТНОШЕНИЕ ДРЕВЕСНЫХ ПОРОД К ЗОЛЬНЫМ ВЕЩЕСТВАМ ПОЧВЫ
Постараемся выяснить, насколько это возможно при современном уровне знания, особенности отношения к зольным веществам наших главнейших древесных пород, памятуя все время, что мы имеем дело с организмами, живущими в естественных условиях, притом в сообществах, при взаимном их влиянии друг на друга. Вторая задача этой главы будет заключаться в выяснении значения добытого материала в жизни леса.
Потребность древесных пород в зольных элементах изучалась до сих пор главным образом путем многочисленных анализов различных частей древесного организма: листьев, мелких ветвей, стволовой части, коры и т. д.; в меньшей степени – другим путем, именно постановкою опытов в сосудах. Относительно этих методов скажем впоследствии несколько больше, а сейчас обратимся к главным результатам, к каким можно свести добытый лесоводами-химиками материал.
1. Если мы будем сравнивать, пользуясь сводными таблицами анализов, количество золы, заключающееся в годичном приросте надземных частей, с одной стороны, луга или поля, а с другой – какого-либо лесного насаждения, то, несмотря на все колебания в числах, мы имеем возможность с полной достоверностью сделать один вывод, а именно тот, что древесная растительность имеет гораздо меньшую потребность в зольных веществах, чем полевые и луговые растения. Нижеследующая таблица (табл. 12) показывает, например, что годичный прирост букового насаждения содержит в 1 1/2 раза, а соснового – в 4 раза менее зольных веществ, чем урожай пшеницы с такой же площади.
Годичный урожай извлекает из почвы с гектара:
Таблица 12
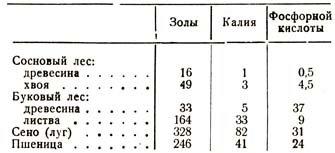
На основании этих и подобных данных мы можем виден,, что лес поглощает из почвы во много раз (и 10 – 15) меньше минеральных веществ, и в особенности таких ценных, как калий и фосфорная кислота. Если мы теперь еще примем во внимание, что он черпает свои зольные вещества, пользуясь большим объемом почвы, чем растения травянистые, то мы будем иметь право сделать другой вывод, что лесная растительность менее требовательна к составу почвы, чем травянистая.
Это обстоятельство объясняет нам факт существования лесов на таких почвах, на которых сельскохозяйственная травянистая растительность удовлетворительно расти не может в силу бедности их зольными веществами. В Западной Европе, в местах густо населенных, как известно, лес постепенно вытеснялся полем и лугом, и эта борьба человека с лесом из-за пространства закончилась оттеснением лесной растительности на так называемые абсолютно лесные земли, т. е. или на такие местности, где в силу рельефа (в горных странах, например) немыслимо возделывать поле, или на пространства хотя и равнинные, но бедные в химическом отношении. Конечно, резкой границы между абсолютно лесными землями и землями, удобными для сельского хозяйства, провести нельзя. Все зависит от экономических условий, от того, в какой мере выгодны те или иные мелиорации, в данном случае удобрения. Еще есть одно следствие двух вышеупомянутых закономерностей, касающееся, правда, уже не леса только, а лесной культуры и лесного хозяйства. Как можно видеть уже из приведенной таблицы и как впоследствии мы это увидим в еще более яркой форме, зольными элементами особенно бедна древесина и, наоборот, относительно богаты листва и мелкие ветви. Но так как главное пользование в лесном хозяйстве есть пользование древесиной, то, значит, вывозится с урожая очень незначительное количество зольных веществ; листва же в виде подстилки остается обычно в лесу. Обратное мы наблюдаем в сельском хозяйстве, где вместе с урожаем поля или луга вывозится не только гораздо более значительное количество золы, но. и наиболее ценная часть ее, именно калий и особенно фосфорная кислота, которая, как известно, находится в почвах в малых количествах. Отсюда – возможность истощения почвы при сельскохозяйственном производстве и невозможность такового, по крайней мере этим путем, для лесного хозяйства, если, конечно, не производится сбора подстилки [1 Академик В. р. Вильяме, опираясь на труды В. В. Докучаева и П. А. Костычева, разработал учение о возможности неограниченного повышения плодородия почвы использованием влияния лесонасаждений, чередованием однолетних и многолетних сельскохозяйственных растений, правильной обработкой почвы, удобрением ее, орошением почвы, что в комплексе с применением отборных семян составляет так называемую травопольную систему земледелия. Ред.]
В качестве иллюстрации приводу еще один пример. Если мы количество калия, которое заключается в годичном приросте сосновой древесины, обозначим 1, то ежегодная потребность в этом веществе елового насаждения будет колебаться около 1 – 2, букового – 2, пшеничного поля – 4, лугового пространства – 10, картофельного поля – 15.
2. Переходим теперь к другим различиям в отношении древесных пород к зольным веществам по сравнению с травянистыми растениями. На основании одного исследования, результаты которого приводятся в табл. 13, можно видеть, что листовая поверхность букового или дубового насаждения значительно меньше, чем листовая поверхность луговой растительности или люцерны (рассчитано «а одном гектаре):
Таблица 13

Но в то же время количество органического вещества, производимого лесом, больше, чем луга или поля. Так, 100-летнее буковое насаждение имеет в виде надземной древесины – 900 000 кг, в виде корневой – 180 000, листовая же поверхность ежегодно – 3 000 кг, а в 100 лет – 300 000 кг, всего – 1 380 000 кг, или ежегодный прирост 13 800 кг, тогда как кормовые травы дают ежегодный урожай только в 4 500 кг. Таким образом, из сопоставления этих данных с прежними, мы можем сделать вывод, что лесная растительность не только принадлежит к менее требовательным элементам в отношении зольных веществ, но еще и производит больше органической массы, или работает более производительно.
3. Если мы далее станем изучать приведенную выше и, приводимые ниже таблицы анализов золы различных частей древесного организма, то в состоянии будем сделать один обобщающий вывод, что чем часть дерева тоньше, тем большее количество золы в ней заключается; так, в среднем количество золы в древесине колеблется около 0,3 – 0,4%, тогда как в мелких ветвях около- 0,8 – 1,2%; богаче всего, однако, золою листья, заключающие в себе 1,3 – 9% ее.
Такая закономерность, конечно, не случайна и находит себе объяснение в том, что в богатых золою частях преобладают живые, деятельные клетки.
4. Благодаря относительно большому количеству золы в листьях л очень малому количеству ее в древесине зольность
Первых элементов послужила лесоводам масштабом для оценки потребности в золе различных древесных пород. Многочисленные анализы показывают, что наименьшим количеством золы отличаются хвойные (1 – 3%), затем береза, ольха и граб, где содержание зольных веществ колеблется около 47% сухого вещества, наибольшим же количеством этих веществ характеризуется ясень и белая акация. Главнейшие породы можно расположить на основании этого критерия приблизительно в следующий ряд, начиная от наиболее богатых золою: белая акация, ильм, ясень, бук, дуб, черная ольха, ель, береза, лиственница, обыкновенная сосна, веймутова сосна.
Как велика может быть разница в величине потребности в зольных веществах у разных пород, хорошо иллюстрирует следующее исследование Раманна. Анализ 30-летней сосны дал следующие результаты (табл. 14):
Таблица 14
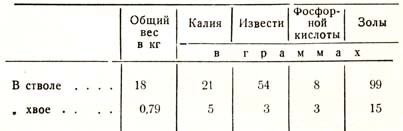
Если приведенные данные для ствола мы разделим на 30, а для хвои на 2Д т. е. на среднюю продолжительность жизни, то получаем следующий ряд данных для годичного прироста сосны (табл. 15):
Таблица 15
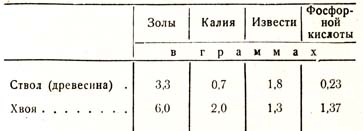
Если мы примем количество зольных веществ, отложенных в годичном приросте сосновой древесины, за единицу, то соответственные данные для хвои будут выражаться следующими величинами:
Золы............1,6
Калия............1,3
Извести.......0,7
Фосфорной кислоты......1,5
Такой же анализ 30-летнего ясеня дал возможность. Раманну получить для тех же веществ в листьях (по отношению к стволу) следующий ряд данных:
Золы...........52
Калия...........22
Извести..........41
Фосфорной кислоты........11
Таким образом, мы видим, что в ту пору, когда сосна тратит на образование листьев, которые притом служат ей 2 – 3 года, 2/3 своей золы, 3л своего калия и 5л своей фосфорной кислоты, – ясень для тех же целей тратит 49/50 всей золы, 21/22 калия и 26/27 фосфорной кислоты. Ясень, согласно этому примеру, для образования своей листвы требует по, сравнению с сосною в 30 раз больше золы, в 8 раз больше калия, в 60 – извести и в 5 раз – фосфорной кислоты. Все остальные породы лежат между этими двумя крайностями.
5. С только что приведенной шкалой пород в отношении их потребности в зольных элементах во многом не совпадают шкалы требовательности к составу почвы, составленные лесоводами на основании наблюдений в природе. Для сравнения привожу шкалы потребности в зольных веществах и требовательности к составу почвы:
Потребность:
белая акация, ильм, ясень, бук,
дуб,
черная ольха,
ель,
береза,
лиственница,
обыкновенная сосна,
веймутова сосна
Требовательность:
ильм,
ясень,
клен,
бук,
граб,
дуб,
черная ольха,
липа,
осина,
ель,
веймутова сосна,
лиственница,
береза,
белая акация,
сосна обыкновенная
Это несовпадение двух шкал происходит оттого, что необходимо различать два существенно разных понятия. Потребность в зольных веществах, определяемая процентом зольности листьев или количества золы в годичном приросте какого-нибудь насаждения с единицы площади, свидетельствует лишь о том количестве минеральных веществ, которое извлекает порода и которое ей необходимо в том или ином возрасте, при тех или иных условиях. Но эта потребность, однако-, еще __ мала говорит об отношении породы к почве, т. е. о большей или меньшей ее способности извлекать нужное количество веществ с тех или Иных почв. Белая акация, например, самая богатая зольными веществами порода, но она, как показывают наблюдения в природе, в состоянии извлекать это большое количество с очень бедных почв. Веймутова же сосна, наоборот, будучи самой бедной по содержанию зольных веществ, нуждается в почвах среднего достоинства и потому оказывается более требовательной к составу почвы, чем обыкновенная сосна. Вот этой-то способностью извлекать нужные вещества из почвы в надлежащих количествах наделены породы в различной степени, отчего и происходит различное отношение разных пород к составу почвы. Эта степень способности мириться с более или менее плодородной почвой и носит название требовательности пород. Белая акация, например, представляет собою пример породы с большой потребностью в минеральных веществах и с малой требовательностью к составу почвы; обыкновенная сосна соединяет в себе малую потребность с малой требовательностью, а ясень – большую потребность, с большей же требовательностью. Различная требовательность пород к составу почвы проистекает от многих причин, которые, однако, изучены еще весьма мало. Прежде всего здесь играет роль способность породы развернуть большую или меньшую корневую поверхность: чем больше будет поверхность деятельной части корневой системы, тем, естественно, в каждый данный момент может поступать большее количество зольных веществ в растение, и наоборот. Можио сказать, что требовательность, при прочих равных условиях, пропорциональна величине корневой поверхности и ее деятельной части в особенности. В качестве иллюстрации сказанного всегда приводятся результаты исследования Ноббе над корневой системой однолетних всходов сосны, ели и пихты (табл. 16):
Таблица 16

Приведенные данные показывают, что число корней у сосны 12 раз превышало число их у ели и почти в 24 - у пихты: общая длина всей корневой поверхности сосны превосходила почти в 6 раз таковую же у ели и в 12 – у пихты; наконец, корневая поверхность у сосны превосходила в 5 раз еловую и в 8 раз – пихтовую.
Приведенные данные вполне подтверждают наблюдения в природе, сделанные в отношении этих трех пород, так как сосна как раз считается наименее требовательной к составу почв породой, за ней следует ель, а затем – пихта. Из своих исследований я могу привести тоже некоторые данные, свидетельствующие о способности различных древесных пород разбивать при равных почвенных условиях различной величины корневую поверхность. Привожу средние величины корневых систем сеянцев, выращенных в сосудах на гумусовом горизонте почвы (табл. 17):
Таблица 17

Эту способность развивать ту или иную корневую поверхность, доказанную для молодого возраста, древесные породы сохраняют и в последующую жизнь, как показывают наблюдения в природе. Помимо большой поверхности, для выяснения требовательности надо обратить внимание на глубину распространения, на тот район почвы и грунта, который в состоянии завоевать та или другая порода. Затем, вероятно, имеет значение характер тех корневых выделений, которые играют роль растворителя почвенных частиц, а также соотношение между корневой поверхностью и поверхностью листовых органов. У одних пород, как, например, у сосны, корневая поверхность характеризуется чрезвычайной пластичностью, дающей ей возможность жить в разных условиях, применяясь к особенностям последних. У других пород, например у ели, нет такой пластичности в этих органах, и потому более ограничен круг тех почвенных сред, на которых они могут произрастать при наличности борьбы за существование с другими породами. Приложенные фотографии (рис. 62, 63, 64) довольно отчетливо, показывают, насколько варьирует корневая система сосны. В одних условиях у нее развивается сильный, глубоко идущий в почву стержневой корень; на болотах у нее, наоборот, стержневой корень не развивается или имеется только в зачаточном виде, а вся система поверхностная. То же самое, если вблизи дневной поверхности находится непроницаемый для воды слой жерствы или ортштеина; тогда стержневой корень развивается слабо, заворачивается в пределах капиллярного подъема воды в стороны, давая разнообразные формы горизонтальных корневых сеток или корневого войлока.

Рис. 62. Корень сосны на слабо развитых дюнных песках (нормальный)
При повреждении стержневого корня ближайшие боковые, подчиняясь геотропизму, невидимому, стремятся его заменить. На почвах очень сухих стержневой корень сосны развивается относительно слабо, но зато сильно развиваются баковые; наоборот, при более близком стоянии грунтовой воды стержневой корень развивается сильно, так что окончание его находится обычно вблизи капиллярного подъема воды. Здесь не место входить в другие подробности, и потому сказанным пока ограничимся.
6. Потребность древесных пород в зольных веществах варьирует затем в зависимости от возраста. Максимум потребности совпадает, как оказывается, с возрастом жердняка, как выражаются в лесоводстве, т. е. с тем возрастом, когда насаждение наиболее энергично растет в высоту, когда оно переживает закон большого периода роста и когда оно характеризуется производством наибольшего количества хвороста, а стало быть и листовой массы.
Нижеследующие данные (табл. 18) для нескольких пород показывают, что есть определенный возраст, когда производство хворостяной массы кульминирует.
Таблица 18
Количество хворостяной массы на один гектар в плотных куб. метрах

В одном из предыдущих пунктов было указано: общее положение, по которому количество золы в разных частях древесного организма повышается с уменьшением диаметра древесины. Исходя из этого и только что приведенного факта, что в жизни каждого лесного сообщества есть определенный период времени, когда хворостяная масса, а стало; быть, и листовая поверхность кульминирует, мы должны сделать вывод, что потребность древесных растений в зольных веществах увеличивается до определенного возраста, достигает максимума, а затем падает.
Изложенное в этом пункте представляет большой интерес для характеристики той лесоводственной точки зрения, о которой шла речь в начале главы. На самом деле, интересуйся мы древесным растением как таковым, без отношения его к общественной жизни, мы едва ли натолкнулись бы на эту закономерность. Дерево, развиваясь на свободе, будет с каждым годом увеличивать свою крону, так что количество хворостяной массы будет безостановочно увеличиваться, в особенности у теневыносливых пород; то небольшое очищение ствола от нижних сучьев, которое будет наблюдаться на свободе, едва ли повлияет на уменьшение хворостяной массы с возрастом. Поэтому и потребность такого древесного изолированного ствола, растущего на свободе, в зольных элементах будет неопределенно увеличиваться с возрастом. Другой ход роста деревьев в лесу, более быстрое и более высокое очищение ствола от сучьев, иное развитие кроны вызывает иное соотношение между развитием хворостяной и стволовой массы и потому влечет за собой перелом в величине потребности в зольных элементах. Тот возраст, в котором эта потребность достигает

Рис. 63. Сосна па болоте
максимума, получил в лесоводстве название критического возраста. Если состав почвы не вполне отвечает данной породе, то обнаруживается это обыкновенно не в ранней молодости, когда может хватить пищевых веществ, а именно в возрасте жердняка, когда вместо сильного прироста последний будет сильно падать и вместо энергичного хода борьбы за существование последняя станет тоже более вялой, ослабеет процесс дифференциации насаждения на господствующие и угнетенные классы.
7. Из относительно малой потребности древесных пород к зольных веществах, конечно, нельзя делать вывода, что химический состав почвы не имеет значения для леса. Многочисленные анализы как почв, с одной стороны, так и древесины, с другой, обнаруживают, во-первых, тот факт, что чем лучше рост леса, тем в большем количестве имеется в почве фосфорной кислоты, калия, извести и пр., а затем, что и. самые растения на лучших почвах отличаются и большей зольностью своих частей. Конечно, не всегда существует такой прямой параллелизм между содержанием зольных веществ в почве и ростом леса, так как в такой сложной среде некоторые факторы могут влиять и в иную сторону.
Рис. 64. Корни сосны у грунтовых вод
8. С лесоводственной точки зрения важно, затем, указать, что зольность одной и той же древесной породы, на одной и той же почве, в пределах одного и того же насаждения тоже будет колебаться в зависимости от того, к какому классу господства принадлежит дерево.
Профессор Раманн, взяв соответствующие деревья – одно из господствующих, другое из согоопюдствующих и третье дерево из угнетенного класса – и определив затем, с одной стороны, прирост деревьев, вес сухого вещества хвои и стволовой массы, с другой – количество; золы, пришел к следующим обобщениям. В среднем 100 г сухой хвои дали в течение 5 лет 206 см3 массы ствола для господствующего дерева, 178 – для согосподствующего и 160 – для заглушённого, или, если выразить продуктивность хвои первого дерева единицей, то таковая же для второго будет – 0,86, а для третьего – 0,78, т. е. разница в усвоении углерода не превышает 0,78 с другой стороны, на 1 г чистой золы приходилось 470 см3 стволовой массы в первом случае, в согосподствующем же дереве – 692, т. е. последняя питалась значительно хуже, чем первая. Выходит так, что сухое вещество угнетенных деревьев отличается меньшим содержанием золы, а отсюда мы вправе заключить, что неодинаковое развитие деревьев разного класса господства находится в связи не только с количеством доступного света, но и с количеством минеральных веществ почвы, могущих быть потребленными теми или иными экземплярами насаждения.
9. Важно отметить, что расчленение питательных веществ, которое делают физиологи, на вещества, безусловно необходимые, и на такие, которые не нужны в искусственных опытах выращивания растений, как известь, кремнезем и пр., – с лесоводственной точки зрения не представляется столь значительным. В жизни растений в естественных условиях – при взаимодействии друг с другом, при необходимости испытывать и влияние животных, и влияние атмосферных деятелей, и т. д. – и известь, и кремнезем, которые откладываются, например, в таких больших количествах в коре некоторых пород, могут иметь большое значение в борьбе с паразитами животного мира и пр. По отношению к таким веществам говорят не о физиологической их необходимости, а об экологической неизбежности.
10. Необходимо отметить затем, что, помимо различения древесных пород по потребностям в зольных элементах, между ними можно различать еще такие, которые очень требовательны к тем или другим веществам; так, для лиственницы характерно высокое содержание магнезии во всех частях дерева, для пихты – калия, для ясеня – извести и т. д.
11. В самое последнее время наши сведения об отношении древесных пород к зольным веществам обогатились новым открытием. По отношению к сельскохозяйственным растениям было давно известно, по отношению же к лесным показано только недавними исследованиями Раманна, что не только различные растения имеют различные потребности в зольных веществах, но; и одни и те же растения изменяют свои требования в разные сроки.
Так, исследованиями названных ученых выяснено, что ель, например, потребляет калия в начале вегетационного периода слабо, около середины июля – сильно, а затем – умеренно до поздней осени; сосна же с самого начала до июля – умеренно, после чего до конца периода – сильно. Фосфорную кислоту пихта воспринимает с начала вегетационного периода до июля умеренно, сосна начинает воспринимать ее только с июля, но зато сильно; ель совсем не воспринимает до начала роста, потом воспринимает умеренно до сентября. Все подобные соотношения по отношению к четырем породам и к трем родам веществ представлены в табл. 19, где посредством вертикальных черточек обозначена степень восприятия вещества, а именно.- I означает слабое восприятие, II – умеренное, III – сильное и IIII – очень сильное; весь вегетационный год, как видно из схемы, разбит на четыре срока.
Таблица 19
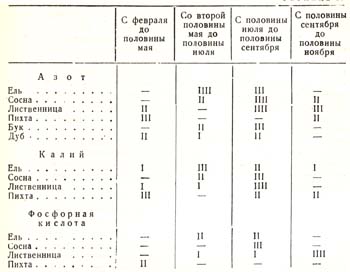
Эти же соотношения показаны и в другой таблице (табл. 20), где слабое восприятие того или иного вещества какой-либо породы изображено простым шрифтом, среднее – курсивом, а сильное – жирным шрифтом.
Таблица 20

Из этих таблиц мы можем сделать вывод, что некоторые породы являются друг для друга в большей мере конкурентами, другие, наоборот, в меньшей. Так, например, пихта и сосна или пихта и лиственница в отношении всех веществ не являются конкурентами друг другу и т. д. Из этих же таблиц следует, что чистые насаждения какой-либо породы предъявляют большие требования к почве, чем смешанные, где те или иные питательные вещества могут быть, по крайней мере при известной комбинации пород, использованы более равномерно; если в насаждении будут смешаны породы, воспринимающие фосфорную кислоту или другое вещество в больших количествах, одни – в один срок, а другие – в другой, то естественно, что они легче покроют свои потребности и будут менее соперничать друг с другом; например, в смешанных насаждениях из пихты и бука первая порода будет воспринимать азот сильно лишь осенью, после опадения листвы у бука, и раннею весною; то же самое в смешанных насаждениях сосны и дуба, где восприятие азота у дуба происходит с весны по июль, у сосны же – сильное восприятие начинается лишь с половины июля.
И этот небольшой очерк отношения древесных пород к составу почвы дает все же некоторую возможность осветить некоторые вопросы из биологии леса. Нам известно из наблюдений в природе, что некоторые породы образуют чистые, другие – смешанные насаждения, а третьи способны образовывать и те и другие. Мы видели также из очерка теневыносливости пород, какое значение имеет в этом отношении фактор светолюбия; теперь остановимся на лесообразовательной роли только что изложенного фактора. Если породы очень мало требовательны к составу почвы, то они в состоянии образовывать сообщества на бедных и мало плодородных грунтах, например сосна на кварцевых песках или на сфагновых болотах. Такие малотребовательные породы, будучи способны образовывать сообщества на бедных грунтах, вместе с тем не встречая конкурентов, образуют в; этих случаях чистые насаждения или с такой незначительной примесью других пород, которая не изменяет сути дела. К сосне, например, на бедных песчаных почвах может быть примешана в небольшом количестве береза и даже ель или дуб, но в виде таких чахлых, еле прозябающих, хронически голодных экземпляров, которые не могут выбиться из стадии чрезвычайно редкого подлеска. Участие в составе насаждения в этих случаях более требовательных пород характеризуется тем, что они не в состоянии сменить собою основное или господствующее насаждение. Наблюдая сильное распространение чистых сосновых насаждений, сделали вывод, что сосна любит песчаные почвы; на самом деле это совсем неверно, так как сосна еще в большей мере любит супеси и легкие суглинки, если судить об этой любви по размерам, каких достигают деревья. На самом деле, на супесях сосна не только развивается пышнее в силу большего плодородия таких почв, но характеризуется и большим плодоношением, образуя здесь насаждения смешанные и сложные.
Лучше всего можно видеть, до какой степени неверно приведенное выше выражение, на примере сосны на болотах; здесь, помимо других причин – избытка влаги, наличности гумусовых кислот, недостаточности кислорода для дыхания корней, – несомненно, играет роль и бедность зольными элементами сфагнового торфа. И сосна, будучи еще в состоянии мириться с этими условиями, находится вместе с тем во власти их, что и отражается на всем габитусе сосны по болоту. Мало требовательные к составу почвы породы – сосна, затем береза – не любят пески и болота, а мирятся с ними. По мере улучшения почвенно-грунтовых условий, как было уже упомянуто, этим малотребовательным породам приходится уже разделять соперничество с другими; в этом случае светолюбив сосны является фактором, способствующим образованию смешанных и сложных насаждений, так как если бы даже территория первоначально была занята только сосной или березой, то в силу светолюбия этих пород их насаждения с возрастом начнут быстро изреживаться и тем давать возможность поселяющимся под их пологом более требовательным и, конечно, более теневыносливым породам разрастаться; последние могут образовать второй ярус, а затем, по мере дальнейшего изреживания верхнего, войти и в первый. Так бывает и с дубом, и с елью при поселении их под пологом сосновых насаждений. Будут ли они образовывать той или иной высоты второй ярус, или в состоянии будут достигнуть верхнего полога сосны, – это будет зависеть от степени плодородия почвы, или, точнее, от степени соответствия состава почвы той породе, которая поселилась под пологом сосны. Чем больше будет это- соответствие, тем лучший рост и лучшее плодоношение будут отличать эти породы, тем лучше будет у них развита крона, тем труднее будет сосновому подросту существовать под их пологом.
Отношение древесных пород к составу почвы, как видим, определяет не только состав и форму насаждений, но и степень способности к конкуренции, или к борьбе за существование, одной породы в отношении другой или других.
По мере улучшения почвенно-грунтовых условий наличность в составе соснового насаждения других пород будет становиться для сосны все более и более угрожающей, и в природе мы можем видеть все переходные формы.
Если породы требовательны к составу почвы, то- тем самым они уже включают в себя свойства образовывать смешанные и сложные насаждения. Будут ли, однако, образованы те или другие, притом в какой мере и в каком именно составе, – будет зависеть уже от других биологических свойств пород. Например, если большая требовательность к составу почв соединяется с очень большой теневыносливостью, то, несмотря на плодородие почвы, на возможность поселения здесь многих пород, насаждения могут преобладать чистые или с небольшой примесью других. Так бывает в некоторых случаях с буком, с европейской пихтой, а также и с елью. Здесь, при образовании насаждений только что названными породами встречаются два биологических свойства, работающие в противоположном направлении при лесообразовании: большая требовательность этих пород к составу почвы влечет за собой необходимость образования смешанных и сложных сообществ, большая же теневыносливость их всегда будет проявлять другую тенденцию. И вот, в результате такой борьбы двух этих начал, в связи еще с другими моментами, о которых речь будет впоследствии, и получаются в природе как чистые насаждения таких теневыносливых и требовательных пород, как пихтарники, ельники, и пр., так и смешанные – елово-пихтово-буковые насаждения, иногда с примесью к ним светолюбивых элементов.
Сказанным тема не исчерпывается. Здесь и не имеется в виду ее исчерпать, а лишь указать на некоторые примеры для характеристики рассмотренных биологических особенностей как факторов лесообразования.
В заключение напомним, что на почвах бедных процесс дифференциации насаждений идет медленнее, так как каждый отдельный индивидуум развивается хуже, чем на более плодородных. Стало быть, в насаждениях той же сосны, борьба между членами сообщества может происходить более энергично и более вяло, в зависимости от почвы, занимаемой породой. Напомним далее, что зольное питание неодинаково у деревьев одной породы и одного насаждения, но принадлежащих к разным классам господства.
Напомню еще и о том обстоятельстве, что разные породы воспринимают главные питательные вещества в разные сроки, что, как мы видели уже выше, может иметь значение момента, ослабляющего конкуренцию при образовании смешанных сообществ. Помимо малой и большой требовательности к составу почвы, некоторые породы отличаются еще исключительными особенностями. Подобно сосне, мирящейся или переносящей, хотя и с уроном для себя, почвы болотные, наш обыкновенный дуб мирится с щелочными солонцами в гораздо большей мере, чем все другие его спутники, и потому он образует в нашей лесостепи на содовых солонцах низкорослые, чистые, одноярусные насаждения. Этот пример особенно поучителен, так как в биологической природе дуба заключены два начала, оба влекущие за собою образование смешанных и сложных насаждений, именно: светолюбие этой породы и ее относительно высокая требовательность к составу почвы. Светолюбие, как мы видели на примере другой породы, а также из предыдущей главы, не препятствует образованию смешанных насаждений, требовательность же к составу почв неизбежно ведет к этому же в естественных условиях природы. Вот потому-то для дуба и считается характерным образование не чистых, а смешанных и сложных сообществ. И как это ни верно по отношению к большинству случаев, необходимо отметить как бы исключение из этого правила: вышеприведенный пример дуба на солонце – пример, возможный лишь потому, что в числе биологических свойств этой породы имеется одна особенность: способность переносить в известных, по крайней мере, концентрациях такие соли, как сода, глауберова соль, или иначе – некоторую щелочность среды.
Перед нами прошли три породы с различным соотношением у них двух биологических свойств – степени теневыносливости и требовательности к составу почв. У дуба оба эти начала направлены в одну сторону, именно в сторону, как мы видели, образования смешанных насаждений; у ели же и у сосны оба эти начала направлены в разные стороны, притом не похоже друг на друга: у ели требовательность к составу почв влечет за собой образование смешанных насаждений, а теневыносливость должна действовать скорее в противоположном направлении; у сосны же, наоборот, светолюбие – фактор, дающий ей возможность образовывать смешанные насаждения, а малая требовательность к составу почв предоставляет ей возможность, несмотря на первое ее качество, в соответствующих, конечно, почвенно-грунтовых условиях; существовать в чистых насаждениях без соперничества других пород.
Малая зольность хвои сосны, малая потребность в зольных веществах, в связи с сильно развитой корневой поверхностью, а также пластичностью последней, дают нам право сделать вывод, что наша обыкновенная сосна – порода очень мало требовательная к составу почв, а такой совершенно обоснованный вывод дает в свою очередь возможность объяснить наличность в природе чистых сосновых насаждений, а в связи с светолюбием – и другое явление: широкое распространение в природе не только чистых боров, но и смешанных сосновых лесов.
Те же данные по отношению к ели – и большая зольность ее хвои, и большее количество хвои и мелких ветвей на дереве по сравнению с сосной, и меньшая корневая поверхность, и меньшая пластичность и глубина ее корневой системы – все это дает полное право вывести, что ель должна быть, как и есть на самом деле, более требовательной к составу почвы породой, чем сосна. Такой вполне обо снованный вывод, исходящий из изучения внутренних биологических свойств породы, дает возможность объяснить факты более сложного порядка, наблюдаемые в природе: приуроченность еловых лесов к суглинкам, к супесям, но ее к перкам, наклонность их к образованию чистых насаждений с небольшой примесью других пород, борьбу сосны с елью и т. д. Такой путь восхождения от простого к сложному – единственно правильный, но, к сожалению, еще слишком мало материала для того, чтобы пользоваться им в широких размерах.
ОТНОШЕНИЕ ДРЕВЕСНЫХ ПОРОД К ВЛАГЕ
Древесные породы нашей флоры принадлежат к растениям, требующим больших количеств влаги для испарения. Есть несколько особенностей у них, которые выделяют их в особую группу. Представители травянистой флоры, произрастая в какой-нибудь засушливой местности, могут бороться с недостатком влаги сокращением своего вегетационного периода: надземные части их отмирают, и жизнь при таких условиях сохраняется в тех или иных формах, чтобы весною, т. е. с новым гражданским годом для растений, проявить себя опять в образовании надземных органов. Древесные породы неспособны к такому сокращению вегетационного периода. Правда, развертывание листьев весною и опадение их осенью у одних бывает несколько ранее, у других позже, но большой, существенной разницы в этом отношении нет. К этому основному различию следует присоединить еще то обстоятельство, что восходящий ток у древесных пород должен быть поднят на большую высоту; необходимо, чтобы на этой высоте отток равнялся притоку, а эта особенность, без сомнения, ставит испарение древесных пород в особые условия. Необходимо затем обратить внимание на то, что древесные породы характеризуются большой листовой поверхностью и произрастают в сообществах. Наряду с условиями, которые повышают испарение и, следовательно, потребность этих растений во влаге, надо обратить внимание на то обстоятельство, что древесным породами свойственна корневая система, более или менее глубоко уходящая в почву и грунт, что не только увеличивает всасывающую поверхность и тот объем грунта, из которого корни могут черпать влагу, но в некоторых случаях благодаря этому делается им доступной и грунтовая влага.
Древесные породы отличаются различной требовательностью к влаге, что запечатлено в различных лесоводственных скалах.
Первоначально породы располагались лесоводами приблизительно в следующий ряд, начиная с пород более требовательных: черная ольха, ясень, клен, дуб, граб, береза, осина, бук, липа, ель, лиственница, пихта, сосна.
По М. К. Турскому, породы можно расположить следующим образом: черная ольха, ясень, клен, бук, граб, вяз, липа, дуб, осина, ель, пихта, лиственница, береза, сосна.
Такого рода классификации являются результатом простых наблюдений в природе и потому характеризуются, во-первых, субъективностью, во-вторых, местным характером, так как чрезвычайно зависят от того круга наблюдений и тех местных условий, в которых их наблюдал автор.
Объективное изучение вопроса об отношении древесных пород к влаге было начато лесоводами давно. В 1852 г. один из лесоводов, занимаясь исследованием потерь в весе отдельных листьев за сутки, получил следующий ряд (в процентах от первоначального веса):
Ольха.......... 43
Дуб ..........37
Бук ..........36
Береза....... 29
Лиственница.........18
Пихта...........12
Сосна......... 11
Ель ..........10
Мы видим, что здесь выступает некоторая закономерность, указывающая на то, что лиственные породы испаряют больше, нежели хвойные, – положение, которое подтвердилось и исследующими исследованиями, более точными в отношении метода. Указанный метод едва ли можно призвать правильным, так как отрезанный лист, конечно, испаряет влагу иначе, чем лист на дереве.
Другой исследователь помещал древесные породы, взятые в лесу и весьма тщательно выкопанные, в сосуды, наполненные водой, причем герметическая укупорка устраняла возможность испарения с водной поверхности. Путем периодического взвешивания определялась убыль в весе, каковая могла произойти только от потери влаги путем физиологического испарения. Эту весовую потерю воды автор относил затем к листовой поверхности соответствующих растений. Отнеся потерю в граммах к одному квадратному метру поверхности листьев, названный ученый получил следующий ряд:
Ольха.........1250
Граб...........290
Сосна...........255
Береза,...........217
Лиственница.........206
Осина.........155
Бук ..........138
Дуб ..........136
Ель ..........106
И этот метод имеет много недостатков; едва ли корневая система этих растений в герметически закупоренном пространстве, наполненном водою, могла работать нормально. Впоследствии, впрочем, названный исследователь усовершенствовал свой метод, устранив герметическую укупорку.
Третий исследователь исходил при изучении этого вопроса из зольности листовых органов, полагая, что между величиной испарения и зольностью листьев должно быть прямое соотношение, т. е. что чем больше древесная порода испаряет воды, тем больше в. ее листовых органах должно остаться зольных элементов. На этом основании названный автор распределил древесные породы в следующие группы, начиная с наиболее сильно испаряющих: к первой группе отнесены породы, в листьях которых заключается золы 7 – 10%, а именно: ясень, ольха, клен, ильмовые, тополя; ко второй группе – менее сильно испаряющие и в листьях содержащие 4 – 5% золы: бук, дуб, граб; к третьей – лиственница, пихта и ель, с зольностью от 2,9 до 3,5%; наконец, к четвертой группе, т. е. к породам, наименее потребляющим влагу, отнесены им обыкновенная и австрийская (черная) сосна, с зольностью 1 – 2%. Такая классификация дает опять-таки лишь самое общее представление, которое можно получить и не прибегая к данным о количестве золы, а путем простого наблюдения в природе. Кроме того, самая база этого метода едва ли бесспорно может быть принята критикой, так как совершенно не выяснено, насколько: процесс накопления зольных веществ в листьях есть результат интенсивности испарения.
Этот же автор пользовался и другими методами, например количеством влаги, которое содержат листья разных пород, и получил при этом такую последовательность пород: тополь (70%), ясень (66%), черная ольха, белая акация (64%), дуб, ильм, липа (63%), береза, клен (62%), осина, бук, граб, полевой клен (57%) и т. д. Нет надобности указывать, что этот метод уже ничего не дает для представления о требовательности пород, так как белая акация, например, – порода, могущая произрастать на относительно сухих почвах, – а также береза согласно этому методу помещаются чрезвычайно высоко. Эбермайер изучал еще отношение пород к влаге по такому способу: сколько дней необходимо, чтобы оторванные листья приняли воздушно-сухое состояние? Оказалось, что, тогда как лиственные породы требуют для этого. 3 – 10 дней (ильмовые, осина), хвойные – от 1 до 3 недель.
Большие исследования были произведены Г. Н. Высоцким. Результаты этих исследований приведены в табл. 21.
Листья вяза и бересклета вследствие тонизны их кутикулы и отсутствия значительного опушения, как замечает автор, теряют влагу наиболее быстро; листья же ильма отличаются, наоборот, значительным опушением, которое ставит эту породу наряду с дубом и ясенем. Кустарники, как видим, а также хвойные породы характеризуются тем, что листья их теряют влагу значительно медленнее.
Таблица 21

Но самые точные опыты над испарением различных древесных пород были произведены на Мариабрунской лесной опытной станции ботаником Гёнелем [1 Опыты Гёнеля в наше время нельзя назвать самыми точными. В нашей стране Л. А. Ивановым разработан метод учета транспирации путем быстрого взвешивания срезанных ветвей, что позволяет изучать транспирацию лесных насаждений всех возрастов со значительной точностью. В то же время метод Гёнеля может быть применен лишь к малолетним растениям, притом растущим не в естественной обстановке, а в горшках. Получаемые этим методом данные не могут быть распространены на естественные леса. Ред.]. Они состояли в том, что ряд древесных пород в возрасте 5 – 6 лет был помещен в особые сосуды с землей, которые в свою очередь помещались в другие, цинковые сосуды для избежания сильного нагревания первых; в последних, герметически закупоривавшихся, имелось по два отверстия: одно – для пропуска стволика, другое – для приливания влаги.