8. Необходимо обратить внимание еще на следующее интересное обстоятельство. Осадки малой силы, благодаря задерживающему влиянию полога, могут и совсем не достигать лесной почвы. Действительно, дождемеры, поставленные под пологом, не всегда обнаруживают осадки, тогда как дождемеры на поляне улавливают их. Благодаря этому засушливый или, точнее говоря, бездождный период может оказаться более продолжительным под лесом, чем вне его. Бюлер приводит такой пример, что тогда как на открытом месте период, в течение которого не было измеримых осадков, продолжался с 23 мая по 1 июня и длился, стало быть, 10 дней, рядом, под столетним буковым лесом, он продолжался от 20 мая по 7 июня, или 20 дней, т. е. был (Вдвое длиннее. В другом году такой же бездождный период захватил на открытом месте 14 дней, под пологом же леса 20 дней, причём был еще приурочен к весеннему периоду, чрезвычайно важному для появления и укрепления всходов.
9. Благодаря задерживательной способности лес имеет определенное защитное влияние как в равнинах, так и горных местностях. В горах опасные ливни благодаря указанной способности легко обращаются в безвредные дожди, чем может предупреждаться снос камней в долине, размыв и снос почвы и тому подобные явления. В равнине смена ельников, например, березою, более пропускающей снег, где он, кроме того, и быстрее тает, может иметь, как показали исследования Н. С. Нестерова под Москвою, решающее влияние на характер водополья, на быстрое его наступление, при отсутствии малейшего изменения в климате.
Таким образом, если мы не исчерпали вопроса о биологическом значении изучаемой нами способности леса, то все ж»-1 вскрыли много сторон, имеющих большое значение в жизни леса, что неоднократно еще будет показано в дальнейшем изложении.
Мы видели, что при наличности одной общей для всех лесов задерживательной способности степень проявления ее варьирует вполне закономерно и обусловливает различные способности и различную жизнь насаждений разного состава, формы, густоты и возраста даже в одном и том же климате. Лес живет, меняется в своем составе, изреживается и т. д., и вместе с этим меняется и количество осадков, которые достигают его почвы.
Наконец, в искусственном изреживании насаждений, в введении подлеска, в создании чистых или смешанных насаждений и т. п. лесной хозяин имеет средства воздействия на один из элементов климата и погоды, т. е. имеет своего рода регулятор, который может увеличивать и уменьшать, смотря по надобности, приток атмосферных осадков к лесной площади.
Таково лесобиологическое и лесоводственное значение вопроса, с которого мы начали рассмотрение претворяющей способности леса.
Количество света под пологом
Перейдем теперь к другому элементу атмосферы, который также сильно преобразуется соединением древесных растений в сообщества, или насаждения. Как известно, общий дневной свет состоит из прямого и рассеянного света при ясном небе, при пасмурном же – конечно, только из последнего. Прямой солнечный свет имеет для древесной растительности второстепенное значение, тогда как рассеянный первостепенное: им определяется положение листьев в кроне дерева, величина листовой поверхности, так называемая мозаика листьев. Рассеянным светом пользуются все листья полога, хотя и в различной степени смотря по глубине расположения их в кронах; прямым же солнечным светом могут пользоваться только листья, расположенные по периферии крон.
Пользование светом в лесу весьма разнообразно: больше всего им пользуются изолированные деревья или смолоду растущие на просторе или потом выставляемые на простор; затем опушечные деревья и экземпляры I класса по Крафту, вершинки которых возвышаются над общим пологом; уже меньшим количеством света пользуются господствующие деревья, образующие верхний полог, и, наконец, деревья угнетенные. Минимум света выпадает на долю нижнего яруса, подлеска, подроста, и, само собой разумеется, почвенного покрова. Если полог насаждения сравнивают в отношении осадков с продырявленным зонтиком, то такое сравнение отчасти применимо и к данному вопросу: часть рассеянного света проникает в промежутки между кронами отдельных деревьев и между листьями кроны, другая часть поступает под полог, уже побывав в листьях после произведенной фотохимической работы. Под пологом насаждений меньше
Света, так как только часть его непосредственно проникает под кроны, а другая часть его поглощается листовой поверхностью, и вот эта-то степень затенения пологом насаждения пространства, заключенного между почвой и пологом леса, будет зависеть от тех самых моментов, от которых зависит и влагозадерживательная способность лесного полога, а именно: от полноты насаждения, от его состава, формы, возраста, климатической области и положения среди леса – опушки, например, могут пользоваться доступом бокового света, физиологически более деятельного, чем верхушечный свет. Исследования, или, вернее, попытки к оным, сделаны были еще в средине прошлого столетия как раз тем методом, который был уточнен Визиером [1 По исследованиям В. Г. Нестерова (см. К вопросу о влиянии леса па снежный покров, журнал «Метеорология и гидрология, 1940 г., № 3), дубовые насаждения разных полнот в Воронежском учебно-опытном лесхозе в 1929 г. в августе месяце пропускали под свои кроны следующее количество света: на открытом месте – 100%, при полноте древостоя 0,3 – 46%, при полноте 0,7 – 12% и при полноте 0,9 – 6%. Ред].
Как известно, венский ботаник предложил измерение количества света, проникающего под полог насаждения, с помощью изобретенного Бунзеном и Роско фотографического метода, основанного на потемнении особым образом приготовленной бумаги. Теоретическим основанием служит тот закон Бунзена и Роско, по которому степень потемнения бумаги прямо пропорциональна интенсивности и продолжительности влияния на нее света. За единицу меры принимается количество света, необходимого для того, чтобы в 1 сек. светочувствительная бумага приняла нормально черный цвет; если последнее будет достигаться в 2 – 3 сек., то интенсивность света будет равна ?, 1/3. Отношение интенсивности света, получаемого растением, к интенсивности общего дневного света (i/y) Визнер назвал фактическим использованием света или величиною использования света; такое же отношение при i = 1 названо им специфическим использованием света, выражающимся всегда правильною дробью с числителем, равным 1, показывающей, какая часть дневного света, падающего в данный момент на данную площадь данного растения, используется.
Этим методом Визнера, дающим лишь представление об относительной, а не абсолютной величине света, да притом еще измеряющим только сильно преломляющиеся, так называемые химические лучи, воспользовалось лесоводство для определения того количества рассеянного света, который проникает сквозь полог насаждения. Проф. Цизляр, пользуясь этим методом, определил количество света под пологом буковых насаждений разной густоты, затем – черной сосны и пихты. Под открытым небом, на поляне, в 40 шагах от букового насаждения, при совершенно ясном небе, 21 апреля, когда еще только единичные экземпляры начали распускать почки, времени для достижения светочувствительной бумагой нормального тона понадобилось в среднем из нескольких наблюдений 4,7 мин; под пологом же букового насаждения, очень сильно прореженного (с удалением почти половины дерев), – 9,8 мин; на следующей пробе, где из насаждения было удалено около А древесного запаса, понадобилось уже 11 мин. и, наконец, в соседнем участке того же леса, где было произведено только обычное сильное прореживание (с удалением IV класса по Крафту), необходимо было 18,2 мин. Если выразить эти величины в интенсивности освещения, то мы получим:
На открытом место....................1/4,7 = 0,213
Под буковым насаждением с сильной рубкой простора…..1/9,8 = 0,102
Обычная рубка простора.................1/11 == 0,090
Сильное прореживание..................1/18,2 = 0,055
Принимая интенсивность освещения под открытым небом равной 1, получим следующие относительные величины интенсивности освещения:
На открытом месте........... 1
Под буковым насаждением:
в первом случае........... 0,48
во втором «........... 0,43
и в третьем «........... 0,26
Такие же наблюдения в июле месяце над насаждениями черной сосны дали следующие результаты:
На открытом месте........... 1
Под рубкой простора.......... 0,42
При сильной проходной рубке........ 0,33
« слабой « « ....... 0,21
Для пихты были получены следующие данные:
На открытом месте........... 1
Под рубкой простора........... 0,24
Под пологом сильной проходной рубки...... 0,10
При слабом прореживании......... 0,07
На основании этих исследований можно сказать, что полог пихтовых насаждений среднего возраста, в зависимости от степени густоты, задерживает от 76 до 90% химических лучей полного освещения; насаждения черной сосны, в зависимости от того же момента, уже гораздо меньше – от 60 до 80% и буковое, ранней весной – от 50 до 75%.
Дальнейшее наблюдение над облиственным буковым насаждением и сравнение его с предыдущим показало, что благодаря листве бука задерживается в два раза больше света, чем в безлиственном состоянии; так, количество света, задержанное кронами бука в зимнем их состоянии, выраженное в процентах от интенсивности света под открытым небом, выражалось в следующих величинах:
Для участка слабо прореженного – 72%
« « умеренно прореженного – 62%
« « сильно прореженного – 56%
В облиственном же состоянии соответствующие цифры таковы: 97% – 96% – 94%, т. е. в 1
1/2 – 2 раза больше.
Приведенные данные подтверждают прежние субъективные наблюдения лесоводов, отмеченные мною уже выше, что количество света, проникающее под полог насаждения, зависит от породы, а следовательно, и состава насаждения, зависит от полноты, т. е. от густоты древостоя и степени сомкнутости крон, а на основании всего вышеприведенного это количество света должно также зависеть от возраста насаждения и его формы. Мы видим также, что теневыносливые породы о густым охвоением или облиствением, как, например, пихта и бук, пропускают под свой полог меньше света, чем светолюбивые породы, как, например, черная сосна, хотя надо заметить, что для типичных светолюбов, как, например, лиственницы, обыкновенной сосны, до сих пор нет наблюдений, произведенных только что- описанным методом.
Ботаник Цедербауер, работавший на Мариабрунской опытной станции, указал на качественную сторону света и решил исследовать свет под пологом леса со спектроскопом в руках. В 1907 г. в печати появилась его работа, которая переведена на русский язык и помещена в 9-м выпуске «Лесного журнала» за тот же 1907 г. Основывается Цедербауер на том положении, что у различных пород, вероятно, различна избирательная способность по отношению к свету, так как, по исследованиям Тимирязева и других исследователей, установлено, что хлорофилл в спиртовом растворе или в живом листе различно поглощает лучи солнечного света. Хлорофилл в спиртовом растворе имеет максимум поглощения между фрауэнгоферовской линией В и С в красной части спектра, около F (голубая часть), J и H поглощение еще довольно сильно, а между С и Е очень слабо. Такую же картину дает спектроскопическое исследование листьев.
Цедербауер старался помещать спектроскоп, как он говорит, в такие места насаждений, где невидимому действие рассеянного света, проникающего прямо под полог, сводилось к нулю. Он производил наблюдения над совершенно сомкнутым пологом ели и бука и получил спектр, похожий на таковой для лучей, прошедших через спиртовую вытяжку хлорофилла, а именно: эти теневыносливые породы, кроме красных, некоторого количества оранжевых лучей, сильно поглощают также, начиная от голубых до ультрафиолетовых. Когда же Цедербауер наблюдал сосну и лиственницу, спектр получился совершенно другого характера: они почти одинаково поглощали в красной, голубой и фиолетовой частях спектра.
Ясень и дуб заняли середину между двумя крайностями а пропускают довольно много лучей в синей части спектра.
Производя в течение двух лет исследования о ручным спектроскопом в целях определения качества света под древесными кронами, Цедербауер нашел, что 1) крона, подобно листьям, обладает избирательной способностью, 2) что поглощательная способность различных древесных пород различна.
Исследования качества света, поглощаемого деревьями, показывают, что деревья предпочитают красные, голубые, фиолетовые и синие лучи, что древесные породы отличаются друг от друга по предъявляемым ими к свету требованиям. Отсюда вытекает, говорит Цедербауер, что при исследовании нужно определять не количество поглощаемых данной породой лучей вообще, а важно знать именно, сколько красных, голубых, зеленых и т. д. лучей она поглощает. Он указывает, между прочим, на неудовлетворительность приборов, с которыми ему приходилось работать, но все же думает, что его данные могут даже претендовать на точность.
Мы видели, что ель, пихта, бук сильно поглощают в красной и от голубой до фиолетовой включительно части спектра, тогда как сосна, лиственница, береза в синей части спектра поглощают мало, а в красной, голубой, фиолетовой частях также менее, чем первые. Сосна, например, растущей под ней ели красных лучей дает мало, но зато пропускает много голубых и фиолетовых лучей, которые и используются елью.
В доказательство целесообразности своего метода Цедербауер приводит, между прочим, указание Борггреве на то, что свет при прохождении через крышу листьев отчасти изменяет свой спектр.
Основываясь на данных своих исследований, что деревья поглощают главным образом красные, синие и фиолетовые лучи, и подчеркивая акт, что при взятии фотографической бумаги галоидные соли серебра будут чувствительны к ультрафиолетовым лучам, Цедербауер указывает на недостаточность метода Визнера.
Влияние леса на температуру воздуха и почвы
Каждый полог насаждения отнимает известное количество тепла, притекающее к его поверхности, но в рассмотрение этого вопроса мы входить не будем, так как имеющийся материал дает общие выводы, давно имеющиеся в курсах общей метеорологии; важные же для нас количественные различия в этом направлении пока еще не разработаны в такой море, чтобы ими можно было реально пользоваться при желании отдать себе отчет в явлениях, наблюдаемых под пологом насаждений хотя бы различного состава. Без всякого сомнения, со временем и этот вопрос будет так изучен, что можно будет привести характерные цифровые выражения для характеристики различия в этом направлении полога еловых, дубовых, сосновых насаждений, простых и сложных форм леса, густых и редких древостоев и т. п. Вопрос же этот весьма важный, так как лесоводственные наблюдения показывают, например, что если для какой-нибудь породы и достаточно еще известное количество тепла, определяемое положением данного места, то подросту данной породы, живущему под материнским пологом, отнимающим известное количество тепла, последнего может уже не хватать, и, по крайней мере, тепло будет отпущено ему, благодаря конкуренции материнского насаждения в такой скупой дозе, что подрост будет представлен в весьма малом числе и весьма угнетенном состоянии. Подрост такой породы, как, например, дуб, на северной окраине или в верхнем пределе своего распространения – в горах может оказаться, благодаря недостатку тепла в таких критических местностях, малоустойчивым, маложивучим под пологом своего же материнского насаждения. Для подроста под пологом насаждения для преждевременной его смерти или для крайнего его угнетения решающими моментами могут быть и недостаток осадков, и недостаток света, и недостаток тепла, причем все эти недостатки проистекают из одного и того же источника – конкурирующего влияния материнского полога. В одном случае решающую роль сыграет недостаток осадков, в другом – недостаток света, в третьем – недостаток тепла; в каждом отдельном случае иногда весьма легко определить, какой из указанных факторов, играет роль закона минимума, иногда, наоборот, их трудно расчленить, тем более, что световая энергия неотделима от тепловой.
Другое явление, которое обращает на себя внимание, – это охлаждающее влияние елового подлеска на такую теплолюбивую породу, как дуб; дурное влияние елового подлеска на верхний дубовый ярус коренится, конечно, в ряде причин, но одна из них, и притом в области распространения дуба, на которую указано было выше, может заключаться в задержании елью притока тепла к почве, занятой дубовым насаждением. Этот вопрос представляет особенный интерес для нашей страны при рассмотрении вопроса о сменах дубовых лесов еловыми, т. е. при изучении вопроса о борьбе дуба с елью, которую покойный Коржинский односторонне поставил в связь с теневыносливостью ели, с одной стороны, и светолюбием дуба – с другой.
Благодаря задержанию известного количества тепла кронами деревьев под пологом наблюдается другой тепловой климат: 1) как известно, почва, занятая лесом, и подрост, на ней живущий, защищены пологом леса как от инсоляции, так и от излучения: лесная почва, как и подрост, получают меньше тепла, но и меньше его излучают; 2) как известно далее, умеряющее влияние леса в большей мере наблюдается в инсоляции и в меньшей степени в отношении излучения; 3) лесная почва в среднем годовом холоднее почвы открытого места, причем, как показали наблюдения, ель влияет сильнее, понижая среднюю годовую температуру на 2 – 3°, тогда как дуб – на 1 1/2 – 2 1/2, а лиственница 0,7 – 2°; 4) известно также, что максимум различия в температуре воздуха и почвы под лесом по сравнению с безлесным пространством наблюдается летом, затем весною и осенью и менее всего зимою; 5) лес повышающим образом влияет на минимумы и понижающим на максимумы; почва под лесом промерзает на меньшую глубину и в меньшей степени; ночью, благодаря защите от излучения, воздух под пологом леса меньше охлаждается, и минимумы не так резки. Благодаря всему этому у деревьев, образующих насаждение, позже пробуждается камбий весной, чем у свободно стоящих; почки у подроста под пологом леса позже распускаются, чем у таких же экземпляров на поляне; у всходов и молодых растений под полотом насаждений большая гармония между оттоком влаги благодаря испарению листьями и притоком в корни, тогда как на открытых местах молодые растения, ничем не защищенные от инсоляции, могут начать транспирировать своими надземными частями тогда, когда корни их находятся или в замерзшей, или в еще очень холодной и потому мало отдающей влагу почве. На открытых местах благодаря отсутствию защитного покрова всходы чувствительных к заморозкам древесных пород обыкновенно побиваются весенними утренниками, так как ничто их не защищает от излучения, с одной стороны, а с другой, благодаря большому притоку тепла они раньше распускают свои почки, часто даже в тот критический период на-начала растительного года, когда бывают еще заморозки; наоборот, такие же растения, но живущие под пологом материнского или какого-либо другого защитного насаждения, будучи защищены от излучения и позже распускаясь, не страдают от заморозков. В природе поэтому породы, боящиеся заморозков, как пихта, ель, бук, дуб и др., обычно не заселяют открытые места, а как бы ждут, пока последние не затянутся так называемыми пионерами древесной растительности – таким быстрорастущими породами, как береза, осина, сосна и др. Когда последние на открытом месте, где велико излучение, из простой совокупности большого множества деревец благодаря наступлению сомкнутости превратятся в сообщество, или насаждение, тогда, только создастся защитный покров, под благодетельным влиянием которого и начнут селиться породы, боящиеся заморозков.
Пользуюсь сим случаем, чтобы показать, что в сложных отношениях, какие устанавливаются между древесными растениями в лесу, наряду с моментами конкуренции, или соперничества, наблюдаются и моменты покровительства защитного характера.
С точки зрения влияния тепла весьма ценный материал могут представить фенологические наблюдения, в особенности если придать им более лесоводственный характер и обратить внимание на время наступления различных фаз в развитии растительности в зависимости не только от экспозиции почвы и тому подобных моментов, но и того, находится ли данное растение под пологом леса или вне его, под каким именно пологом леса – ординарным, двойным или тройным, под густым или редким, под пологом той или иной породы, того или иного возраста и т. д.
Фенологические же наблюдения должны показать, что вегетационный период под пологом леса, вероятно, короче, чем на открытом месте.
По сообщению С. Д. Охлябинина, например, раскрывание шишек сосны в 1909 г. в Бузулукском бору на открытых местах началось 21 апреля, а в лесу только 2 мая, в первом случае на 26-й день от начала весны (т. е. от начала таяния снега), а в лесу на 36-й день, причем на это потребовалась сумма температур: на поляне 60,1°, а в лесу 143,3°.
Принимая во внимание, что под пологом леса температура воздуха ниже, а амплитуды меньше, проф. Ра манн говорит, что облесение какой-либо поверхности земли как бы переносит ее несколько к северу, с одной стороны, и к западу – с другой. Заканчивая этот отдел, нельзя опять не обратить внимания на то, что приток тепла к почве, подросту и составляющим насаждение деревьям можно регулировать так же, как и приток осадков и света, притом и в ту и другую сторону, т. е. и в сторону плюса и в сторону минуса, путем искусственного изреживания насаждения или путем сгущения пологов, введением второго яруса или подлеска. Но кроме такого искусственного регулирования, которое находится во власти хозяина, насаждение само, процессом своей жизни, одной из характернейших черт совместной жизни древесных растений, именно процессом естественного изреживания или самоизреживания, способно изменять приток тепла. Сомкнутая чаща больше всего задерживает тепла; в возрасте жердняка, когда процесс дифференциации насаждения на господствующие и угнетенные классы в самом разгаре, приток тепла усиливается, становясь еще большим в период сильного изреживания, которое наступает сейчас же после кульминации роста в высоту; по мере приближения к возрасту возмужалости насаждение еще более изреживается, что является биологически целесообразным, потому что деревья теперь нуждаются в большом притоке тепла (света и осадков) для проявления своей физической зрелости, для плодоношения. В спелом лесу, окончательно возмужалом, больше всего притекает света и тепла как в области кроя, так и под полог насаждения – к самой почве, на которой теперь уже заметен результат половой возмужалости насаждения, именно – наличность подроста. По мере естественного изреживания насаждения, как видим, улучшаются условия жизни подроста с двух сторон: с одной – благодаря увеличению тепла, света и влаги, с другой – все меньшей и меньшей конкуренции со стороны материнских организмов, так как в долгой борьбе за существование очень значительная часть – около 80 – 90% от первоначального населения – пала жертвою борьбы за существование; да и IB наличном составе деревьев приспевающего и спелого возраста не все плодоносят, как мы видели уже, г. только победители в жизненной борьбе – преимущественно I и II классы по Крафту.
Благодаря защитному влиянию полога на тепловые условия атмосферного пространства, под ним находящегося, под пологом леса, как известно, наблюдается большая относительная влажность воздуха, чем в поле. По наблюдениям Н. П. Адамова и моим в нашем лесостепном районе разность в этом отношении между сухим сосновым бором и соответствующей поляной незначительна и даже непостоянна, тогда как в сложном насаждении, в дубравах Шилова леса, разность в пользу леса, во-первых, – величина постоянная, во-вторых, – гораздо более значительная, достигая среди дня даже 10°.
Аналогичные же данные приводятся для Эберсвальде, где констатируют незначительные разницы для соснового леса и более значительные для букового. Разработка этого вопроси, в зависимости от состава, «формы, густоты, возраста насаждения и нахождения его в том или другом климате, тоже еще находится в зачаточном состоянии и ждет своего исследователя; общие же положения хорошо известны из курсов общей метеорологии.
Отношение насаждения к движению воздуха
Этот вопрос возник вместе с возникновением лесоводства. Отчего же он всегда так занимал лесоводов?
Какое значение имеет ветер для объекта лесоводства – леса?
При виде огромного, высокорослого дерева, с широкой, развесистой, если дерево стоит одиноко, кроной, невольно приходит мысль о той громадной силе, которую приходится выносить дереву под напором движущегося воздуха; но дерево, всегда росшее на открытом месте, так приспособило свою крону, что легко переносит без вреда для себя очень сильные ветры. Но ведь лесоводы имеют дело с лесом, с сообществом деревьев, в котором, если оно не подвержено хозяйству человека, действию ветра подвержены только опушки, внутри же леса, как мы увидим ниже, движение сводится почти к нулю; следовательно, и приспособлены переносить ветер только опушечные деревья. Внутри насаждения деревья, не зная ветра, не могут приобрести форму, выгодную для сопротивления ему, а поэтому, как только появляются лесосеки, то там, то сям внезапно открывающие доступ ветру в насаждение, деревья нередко и а больших расстояниях падают, вываливаются с корнями.
Вот уже то, что деревья высоки, что у них высоко прикреплена крона и они, стало быть, являются большим препятствием для ветра, что деревья растут в лесу, – заставляет лесовода так внимательно относиться к действию ветра на насаждения.
Прежде всего бросается в глаза каждого, видевшего лес, механическое действие ветра на лес. Ветер с корнями выворачивает огромные деревья или ломает их, производя так называемые ветровалы и буреломы.
В насаждении, как мы уже видели, деревья как бы отвыкают от ветра и слабее укоренены, чем на просторе, – и вот при внезапном выставлении насаждения на свободу деревья будут страдать от ветровала. Конечно, эта ветровальность зависит не только от ветра и вообще от (внешних условий, но и от внутренних свойств леса, от способности укоряться в более или менее глубоких слоях почвы. Классической по вертикальности породой считают ель, затем следуют буки пихта. На противоположном полюсе да отношению к ветру стоит дуб.
Далее, влияние ветра сказывается на росте и форме дерева. На прибрежьях морей, где ветры дуют постоянно в одном направлении, деревья растут обыкновенно наклонно, не имеют совсем ветвей на подветренной стороне, наиболее утолщаются по диаметру, совпадающему с направлением ветра. На высоких плато и вообще там, где ветры дуют очень сильно, леса или совершенно нет, или он принимает форму кустарниковых зарослей. Чем более свободно растет дерево, тем более оно подвергается действию ветра, тем оно сбежистее, тем ниже подвешена его крона. Изучая значение различной высоты места произрастания на рост и форму дерева, можно заметить, что деревья тем ниже ростом и сбежистее, чем выше над уровнем моря растут, что, конечно, частично объясняется действием ветра.
Как показали исследования, форма деревьев насаждений, в отличие от форм изолированных их собратьев, может быть сведена на влияние ветра. Благодаря большому весу кроны дерена ствол должен обнаруживать сильное сопротивление излому; и на самом деле, на основании весьма интересных аргументов и наблюдений можно утверждать, что дерево вырабатывает при наименьшей затрате строительного материала форму, хорош сопротивляющуюся изгибу. Древесные стволы, в особенности еловые, наиболее близко подходят под понятие равномерно сопротивляющихся тел. Диаметр такого тела равного сопротивления, при экономии в материале, должен возрастать от свободного конца его к прикрепленному и тем скорее, чем больше нагрузка первого; при большой сгибающей силе, т. е. при большой кроне и свободном стоянии, форма такого сопротивляющегося тела и должна напоминать форму сбежистого ствола; наоборот, при малой силе ветра и малой величине кроны; что мы на самом деле и наблюдаем в насаждениях, форма ствола может стать более полнодревесном. И на самом деле, все эмпирические наблюдения подтверждают теоретический вывод, так как нам хорошо известны: 1) большая сбежистость стволов свободного стояния: 2) увеличение таковой, или понижение видового числа, при переходе деревьев из сомкнутого состояния в свободное, когда происходит под влиянием прироста свободного стояния, или светового прироста, сильное утолщение в нижних частях таких свободно стоящих деревьев, 3) наконец, уменьшение полнодревесности стволов с повышением местности над уровнем моря; наоборот, с ослаблением влияния ветра в сомкнутых насаждениях, в особенности при наличии второго яруса, прирост откладывается более равномерно вдоль ствола; -в последнем случае нет такого значительного увеличения его у основания дерева, и, таким образом, благодаря механическому влиянию ветра, стволы деревьев в этих условиях становятся более полнодревесными. Условия питания определяют величину прироста, ветер же диктует, как этот прирост должен отложиться вдоль ствола. Опушечные деревья, вырастающие смолоду под постоянным напором ветра, и приобретают соответственную выгодную им для сопротивления форму. Совсем обратное происходит с теми деревьями или стенами леса, которые выросли среди насаждения и потому, мало испытывая механическое влияние ветра, создали особые формы: тонкий и голый ствол с высокоприкрепленной кроной; такие отвыкшие от ветра деревья легко становятся добычею ветра; надо добавить еще, что такие деревья и слабее укоренены, чем те, которые смолоду произрастают на просторе.
Ветер усиливает как физическое, так и физиологическое испарения. Ботаники и географы объясняют им карликовый рост деревьев тундр и отсутствие ветвей с наветренной стороны. И действительно, в тундре мы видим, что то, что заносится снегом или растет в логах, может существовать, не закрытые же снегом высокие растения не в состоянии выносить иссушающего Действия ветра.
Ветер имеет большое значение и в биологии леса: почти вес лесные растения, составляющие первый верхний ярус насаждений, ветроопыляемы, и все разносят сноп семена при помощи ветра. Некоторые по особенностям распространения семян делят все породы на три группы: первые – плоды вскрываются и семена разносятся при сухих восточных ветрах – сосна, пихта, ель; вторые – семена открываются и разносятся порывистыми западными ветрами – ясень, липа, клен, и третьи – не зависят от ветра – дуб, бук.
Ветер еще имеет то значение, что производит охлестывание деревьев; III и IV классы по Крафту являются охлестывателями первых двух классов. Особенно заметно вредное влияние охлестывания в лесах смешанных, например ель с березой, – последняя сильно повреждает ель и нередко в борьбе за существование одерживает над ней победу.
Ветер, раскачивая деревья, обрывая корни и тем ослабляя их, подготовляет почву для массового размножения вредных насекомых, например короедов, являясь, таким образом, первичным врагом.
Внутри леса, как мы уже знаем, ветер почти отсутствует и это ведет за собою малую потребность во влаге подроста.
Как видно из предыдущего, ветер имеет огромное, всестороннее значение для леса, и Майр сделал такие выводы:
ветер – создатель леса,
ветер – разрушитель леса,
ветер – препятствователь распространению леса.
Этим последним он хочет сказать о том, что северные границы лесов и границы их на горах обусловливаются ветром.
В своем лесоводстве («Waldbau auf naturgesetzlicher Grund-lage») Майр показывает, что леса Северной Америки на Береговых, Каскадных, Голубых и Скалистых горах в своем распространении зависят главным образом от ветра в связи с его влажностью. Мы видим, что там, где горы омываются ветром, идущим прямо от моря и потому влажным, растут леса; где этого нет – там степи. Отсюда вывод: океаны – создатели леса.
О взаимоотношении между ветром и насаждением есть научно произведенная работа – это работа проф. Нестерова, напечатанная в № 8 и 9 «Лесопромышленного Вестника» за 1908 г. под заглавием «О влиянии леса на силу и направление ветра».
Наблюдения производились автором в даче б. Петровско-Разумовской академии (Сельскохозяйственного института) с ручными анемометрами Робинсона. Наблюдения велись одновременно в различных пунктах в лесу и вне леса, причем пункты эти в большинстве случаев выбирались по линии, соответствовавшей направлению ветра. Лишь в некоторых случаях измерение скорости ветра производилось разновременно, притом с таким расчетом, чтобы через оба анемометра, при действии их. проходила по возможности одна и та же воздушная волна. Во время наблюдений один анемометр находился вне леса на постоянном пункт-е, а другой постепенно перемещался по намеченной линии с одного пункта на другой; при большом же протяжении наблюдательной линии передвигались оба прибора с расчетом, чтобы можно было связать результаты наблюдений. Скорость ветра на каждом пункте отмечалась троекратно, причем анемометр открывался каждый раз на 2 минуты, а при дальнейших расстояниях между пунктами – на 5 минут. Вычислялась средняя из этих данных скорость ветра и выражалась затем в процентах от скорости ветра на открытом месте. Направление ветра определялось по движению дыма от ожигания Polyporus tomentarius (березовый трутовик); иногда автор пользовался даже дымом от папиросы.
Наблюдения показали, что при движении с открытого места в лес скорость ветра близ опушки (начиная примерно с 64 м расстояния) значительно увеличивается против первоначальной скорости, возрастая на 20 – 30% и до 60%, в зависимости от густоты леса, высоты его, наличности нижнего яруса и самой скорости ветра. Кроме того близ опушки, леса нередко наблюдается волнение воздушных течений, подобное прибою морских волн, выражающееся в анемометре в быстрых и частых сменах скоростей, а также в беспорядочных движениях дыма.
Эти воздушные волны повышенной скорости врываются и в самый лес на то или иное расстояние, в зависимости от тех же условий, которые только что упомянуты (в зимнее время расстояние это наглядно выражается шириною снежного наноса на опушке).
Нижний ярус насаждения имеет при этом большое значение как стена, он задерживает движение ветра в нижних слоях. По наблюдениям в 53 м от опушки леса при редком подросте скорость ветра в лесу составляла от 73 до 127% сравнительно о открытым местом, тогда как в 75 м, т. е. лишь на 22 м глубже в лес, среди орешника средней густоты скорость того же ветра на протяжении этих 22 м быстро падала до 17 – 19% (рис. 74).
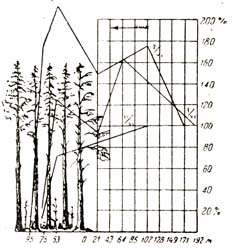
Рис. 74 Скорость ветра, дующего с открытого места в лес

Следующие данные, относящиеся к насаждению из сосны с густым дубовым подростом и орешниковым подлеском, показывают постепенность затихания ветра по мере удаления от опушки.
Куда же деваются те воздушные потоки, которые с громадною силою врываются в лес? Часть их на окраине леса под напором спруженного на опушке воздуха поднимается вверх между кронами, растекается между ними или проходит над лесом. Далее живая сила ветра расходуется в лесу: 1) на трение о стволы и ветви, превращаясь в теплоту; 2) на механическую работу по раскачиванию стволов, движению ветвей и листьев, которая тоже переходит в теплоту, и 3) на работу го обиванию сухих сучьев, молодых побегов, листьев и т. д.
Таким образом в лесу должен был бы царить полный штиль, если бы не было движения воздуха от прогалин, дорог и просек.
Предел проникновения ветра в насаждение особенно рельефно выступает во время изморози, которою покрываются лишь вершины деревьев, что и показывает, что и сверху ветер имеет небольшой доступ в полное насаждение.
Как мы видим, приближаясь к лесу, ветер развивает наибольшую силу, и это ставит опушечные деревья в исключительно невыгодное положение; они, подобно скалам, вечно омываемым морскою волною, подвергаются сильному напору воздуха и притом – как из леса, так и с открытого места. Это обстоятельство, возникающее неизбежно при закладке сплошных лесосек, подвергает существование окраинных деревьев большому риску: разрыв корней и вывалка, плохой рост и засыхание от физиологического недостатка влаги. Недостаток влаги обусловливается усиленною транспирациею их; на этот факт нам часто намекают: засыхание ели, суховершинность березы и дуба, плохой вид сосны, отсутствие возобновления близ опушки и другие факты.
Более всего задерживается ветер в еловых насаждениях. В них царит, можно сказать, постоянное затишье. Так, когда на открытом месте ветер движется со скоростью 374 – 4 км в час, в ельничке 35 – 38-летнего возраста движение воздуха составляет лишь 340 м в 1 час. Такою же ветрозадерживающею способностью обладают и полные сосновые жердняки.
В наименьшей степени задерживается ветер редкими сосновыми насаждениями, лишенными нижнего яруса. Когда ветер дует по направлению к редкому однояруснику, то он при приближении к лесу и у самой опушки немного лишь теряет в своей скорости (2 – 6%) и пропускается далеко вглубь; так, в расстоянии 65 – 75 м от опушки в таком лесу скорость ветра понижается лишь на 5 – 6%, тогда как в двухъяруснике на этом расстоянии, как мы видели, теряется 4/3 силы ветра. 212
Подчеркнем здесь еще раз то огромное лесоводственное значение, какое имеет «тихая» обстановка под пологом леса и связанная с нею малая транспирация растений и испаряемость с поверхности лесной почвы.
Ветер, дующий из насаждения на открытое место, дает нам подобную же картину: чем ближе к опушке, тем он сильнее.
Насколько далеко распространяется влияние леса на окружающую местность, проф. Нестеров не решается отвечать, и только в пределах 850 м от леса он утверждает заметное влияние леса как на ветер (его силу и отчасти направление), так и на испарение воды в эвапарометрах.
Из приведенных выше данных (более подробно с ними познакомиться можно в указанной выше статье) проф. Нестеров выводит следующие заключения:
1. «Ветер, встречая на своем пути такое препятствие, как лес, каждый раз стремится принять направление наименьшего сопротивления, причем, более или менее значительно уклоняясь от своего основного пути, он несется, образуя невидимые вихри, по дорогам и просекам, врывается в редины, прогалины, а также огибает целые массивы леса; в связи с тем на подветренной стороне леса, в известном расстоянии от него, происходят более или менее сложные сочетания сил воздухопада, окольных ветров и основного течения».
2. «Врываясь в насаждения, ветер быстро теряет почти всю свою силу и изменяет направление; при этом существенное значение имеют нижний ярус и полнота насаждения. В нормальных хвойных насаждениях обычно царит почти полный штиль».
3. «Ввиду большой ветрозадерживающей способности леса следует при эксплоатации его обращать особенное внимание на ветроопасные пункты в даче. Здесь необходимо сохранение в целости пограничной опушки, открытой доступу господствующих ветров, а также соответствующая закладка лесосек».
4. «Плохой рост, суховершинность или засыхание опушечных деревьев на сплошных лесосеках являются следствием физиологической сухости, вызываемой усиленным действием ветра на эти деревья».[ 1 Исследования влияния леса на ветер в степных условиях СССР произведены рядом лесомелиораторов. Ред.]
Мы видим, таким образом, что ветер является одним из первостепеннейших факторов в жизни леса, так что по важности он стоит наряду со светом и влагой.[ 2 Ветрозащитное действие леса, так же как его умеряющее влияние на температуры соседних площадей, собирательное действие в отношении снега и другие ценные свойства, позволили в СССР широко развернуть полезащитные лесонасаждения в степных и лесостепных районах для борьбы с сухими юго-восточными ветрами и с засухами с целью обеспечения высоких и устойчивых урожаев хлеба. 20 октября 1948 г. было опубликовано историческое постановление Совета Министров СССР и ЦК ВКП(б) «О плане полезащитных лесонасаждений, внедрения травопольных севооборотов, строительства прудов и водоемов для обеспечения высоких и устойчивых урожаев в степных и лесостепных районах европейской части LLCF», согласно которому в ближайшие 15 лет должны быть созданы восемь государственных защитных лесных полос, каждая длиною в несколько сот и даже в отдельных случаях более тысячи километров, а также полезащитные лесные полосы площадью около 6 млн га Это постановление является сталинским планом преобразования природы степей и лесостепей в целом. Ред.]
Итак, ветер так же, как свет, тепло и влага, как и прочие факторы жизни, действует в лесу постоянно и имеет поэтому прежде всего общее значение. К этому общему значению относится прежде всего его механическое влияние и опять-таки не в виде только ветровала и бурелома, а в виде постоянно действующего раздражителя, под влиянием которого происходи г распределение прироста вдоль по стволу, который лепит из данного материала ту или иную форму ствола в зависимости о; условий, в каких дерево находится.
Вторым общим его влиянием является роль в испарении влаги со стороны почвы и растений; сильное ослабление ветра под пологом леса сильно уменьшает физическое испарение влаги из почвы и физиологическое из подроста деревьев нижних ярусов, а отчасти также верхнего полога. Влияя таким образом на рост и форму деревьев, на испарение влаги в лесу, ветер обусловливает опыление ветроопыляемых деревьев, обычно составляющих первый ярус, разносит плоды, обусловливает вылет семян, а будучи ослаблен под пологом леса, способствует более равномерному залеганию снега, большей рыхлости снежной пелены, что выгодно может отзываться на сохранении лесной подстилки. В насаждении он играет еще одну роль в борьбе древесных растений одной и той же породы и разных пород друг с другом благодаря ему происходит так называемое взаимное охлестывание дерев; деревья, которые легче приводятся в движение ветром, охлестывают соседние и лишают, таким образом, части рабочих зеленых органов растения; особенно популярно в лесоводстве охлестывание березой сосны.
Испарение под пологом леса
Под влиянием уменьшения пологом леса инсоляции под ним, благодаря более низкой температуре, атмосферы под кронами деревьев насаждения, благодаря уменьшению силы ветра и более высокой относительной влажности того же воздуха, условия испарения и величина испаряемости под пологом леса иные и гораздо более благоприятные, чем на открытом месте. Это справедливо по отношению к любому предмету, который может испарять воду и который помещен под пологом леса; еще в большей мере это верно по отношению к тем предметам, которые коренятся в почве, занятой лесом, как, например, подрост, так как лесная почва, испытывая все указанные выше понижающие испарения условия, находится в то же время под защитою такого покрова, как лесная подстилка, которая вместе с другим защитным покровом, именно пологом леса, как это мы видели уже, в свою очередь защищает почву от инсоляции; благодаря тому, что лесная почва более холодна, поступление из нее влаги в корни растений должно быть замедленным. То же обстоятельство, что почва покрыта подстилкою, влияет в одном случае благоприятно, уменьшая испарение с поверхности почвы, сама же подстилка, отличаясь горизонтальной слоистостью испаряет мало; в другом случае, когда подстилка большей мощности и плотности, она, наоборот влияет неблагоприятно на влажность почвы: с одной стороны, она не пропускает часть атмосферной влаги, которая в виде луж долго сохраняется в пониженных участках, пока вся не испарится в воздух; с другой стороны, такая подстилка, благодаря своей массе и большей гигроскопичности, удерживает в себе много влаги. Иногда после сильных дождей почва в лесу бывает так же суха, как и до выпадения осадков, и если снять плотную, кислую подстилку на каком-нибудь пространстве, то почва обнаруживает своеобразный рисунок, где темные пятна и расплывчатые линии чередуются и переплетаются с более световыми местами; там, где были трещины в слое кислой подстилки, по тем местам проникла вода, создав потные места и своеобразный рисунок; там же, где никаких трещин не было, вода не проникла, и почва осталась так же суха, как и раньше.
Но как бы там ни было, если мы будем рассматривать вопрос в общей форме, то физическое испарение влаги из почвы под пологом леса сильно понижено, – по известным опытам Эбермайера, на 62% по сравнению с почвою вне леса, если при этом она не снабжена подстилкой; если же, как это обычно и бывает, почва защищена не только пологом леса, но и подстилкою, то испарение с ее поверхности уменьшается еще на 23%, или всего на 85%. Иначе говоря, если почва в определенный период времени испаряет 100 единиц, то такая же почва под пологом леса, но без подстилки теряет 38 единиц, а прикрытая подстилкою всего только 5 единиц. Из этого факта, к которому возможно присоединить данные других исследований, например Бюлера, можно сделать вывод о том, как велико значение соединения древесных растений в насаждение для уменьшения непроизводительного расхода из почвы влаги. Мы можем далее сделать другой вывод, что создание лесом своеобразных условий под своим пологом должно сильно понижать и физиологическое испарение со стороны подроста, подлеска, затем второго яруса, угнетенных деревьев и даже деревьев господствующих, так как нижняя часть их кроны находится в условиях пониженного Испарения.
Влияние леса на влажность почвы
Для того чтобы в полном объеме представить себе гидрологические условия, в каких находится хотя бы подрост, – это будущее поколение леса, идущее на смену старому, – необходимо отдать себе отчет во всестороннем влиянии насаждения на влажность почвы и грунта, занятого лесом. Исследования этого рода были произведены Эбермайером, причем он пользовался не методом прямых определений влажности, а судил о ее влажности по количеству воды, просачивающейся в течение вегетационного времени сквозь почву, с одной стороны, находящуюся под пологом насаждения, с другой – свободную от леса. Эбермайер пользовался лизиметрами с двойным дном, одним сплошным в виде воронки с одним центральным отверстием, другим, выше расположенным, – дырчатым; эти цилиндрические сосуды были наполнены почвой и закопаны в почву под лесом и вне леса так, что верхний край цилиндра приходился в уровень с поверхностью почвы. Вода, падавшая при дождях на почву в лизиметрах, насытив ее, могла затем стекать из сосудов через отверстия в воронковидном дне так, что ее можно было собирать и измерять. Все наблюдения дали одинаковый результат, а именно, что в лесу сквозь лизиметр просачивается больше воды, чем через лизиметр на поляне; в первом случае через слон почвы глубиной в 0,6 м просочилось 47 539 см3 и вне леса 36 050 см3; через слон почвы в 1,2 м – в первом случае 129 834 см3, во втором 127 360 см3; отсюда Эбермайером, а затем и другими, был сделан парадоксальный вывод, что лес способствует сохранению влажности почвы. Такой вывод, казалось, был совершенно логичен, так как и на самом деле большее просачивание воды под лесом могло быть только при большей влажности лесной почвы. Цифровые данные Эбермайера не подлежат ни малейшему сомнению, а между тем его выводы совершенно неправильны, на что впервые указал у нас проф. Костычев, а затем и сам Эбермайер имел мужество признаться в неверности своих выводов. «Подобные результаты неизбежно должны были получиться при таких наблюдениях, – говорит проф. Костычев, – так как ведь почва, находившаяся в сосудах, была свободна от растений; поставленная под покровом леса, она защищалась от высыхания отенением, но нисколько не иссушалась действием древесных корней; это была, следовательно, совсем не лесная почва, и по ее высыханию нельзя судить о высыхании настоящей лесной почвы». В связи с этим следует упомянуть исследование Ризлера в Швейцарии, который, пользуясь методом непосредственных определений влажности почвы, получил, между прочим, следующие результаты. 25 августа 1869 г. содержалось воды в процентах (табл. 30):
Таблица 30

«Из этих цифр видно, – говорит Костычев, – что лесная почва была суше полевой и особенно высохла в более глубоком слое соответственно распространению корней».
Первое исследование подобного же рода, произведенное в России, притом в гораздо более широком масштабе, принадлежит Вермишеву, тогда студенту Лесного института, который работал под непосредственным руководством профессора П. А. Костычева. Работа его опубликована в 1884 г. и подтвердила результаты Ризлера. Из весьма многочисленных данных, собранных с большой тщательностью, я приведу лишь следующий ряд цифр, тоже относящийся к августу месяцу, в процентах (табл. 31):
Таблица 31
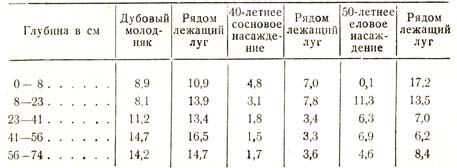
«Цифры эти показывают, – говорит П. А. Костычев, – что деревья уменьшают содержание влаги в почве, и даже, повиди-мому, сильнее травянистых растений в той же почве».
Впоследствии Эбермаиер (1884 г.) снова вернулся к исследованию этого вопроса, но работал уже методом непосредственны.-: определений влажности почвы, причем получил, как это предсказал П. А. Костычев, те же результаты, как и его предшественники – Ризлер, Костычев-Вермишев. В последней и притом сводной своей работе Эбермайер пишет следующее: «Вопрос относительно влияния леса на влажность почвы начал выясняться лишь в последние десятилетия, причем оказалось, что Почва под лесом тем суше, чем больше задерживается влага кронами дерев, чем совершеннее развитие корней и крон и чем дольше вегетационный период». Уже первые наблюдения Эбермайера, произведенные близ Мюнхена, дали ему право высказать следующие положения: 1) верхний слой почвы в лесу, в котором корни не распространены, отличается в течение всего года большей влажностью, чем соответственный слой в поле, что объясняется более слабым испарением на поверхности затененной почвы в лесу, нежели на открытом для солнца и ветра поле; 2) совершенно иное распределение влаги встречаем мы в лесу в более глубоких слоях почвы: уже на глубине 15 – 20 см, а еще более на 30 – 80, в которой находится наибольшая масса корней, лесная почва в течение почти всего года значительно суше, чем на соответствующих глубинах в поле, причем наибольшее количество воды отнимают от почвы жердняки, затем молодняки, а менее всего иссушивают почву старые изреженные насаждения; 3) летом и осенью, в течение наиболее продуктивной деятельности растений, влияние леса на высушивание почвы в глубоких слоях значительно больше, чем зимою и весною. Наиболее существенные результаты исследований Эбермайера сведены им в следующей таблице (табл. 32):
Таблица 32
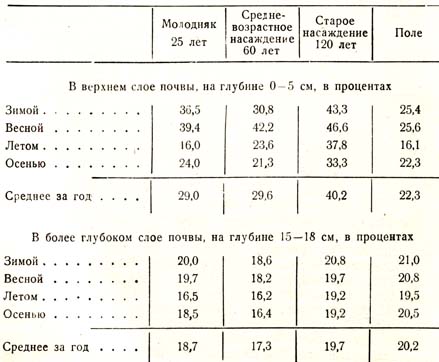
Для выяснения влияния различных классов возраста на степень высушивания почвы Эбермайер приводит следующую таблицу, в которой даны разности в содержании влаги в процентах между полем и еловым лесом трех возрастов по временам года (табл. 33):
Таблица 33
Приведенные числа не требуют дальнейших объяснений.
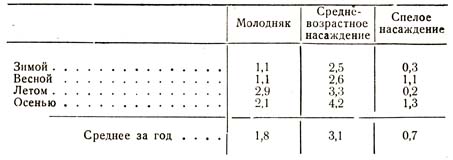
Многочисленные исследования были произведены также проф. Раманном, из числа которых я ограничусь приведением следующей таблицы (табл. 34):
Таблица 34
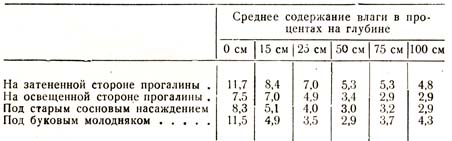
Результаты, полученные проф. Раманном, находятся, как и вообще все исследования этого рода, в полном согласии друг с другом, т. е. решительно все они подтверждают, что влажность почвы в области распространения корней в лесу, в насаждениях любой породы, любого возраста, меньше, чем в тех же слоях безлесного пространства. Глубина, на протяжении которой лес высушивает грунт, больше в старом сосновом насаждении, благодаря более глубокой корневой системе этой породы, чем в буковом, корни которого стелются более поверхностно и высушивают поэтому преимущественно верхние слои.
Наиболее многочисленные исследования по вопросу о влиянии леса на влажность почвы и грунта производил у нас в России Г. Н. Высоцкий в Велико-Анадоле, причем он получил следующие данные (табл. 35):
Таблица 35
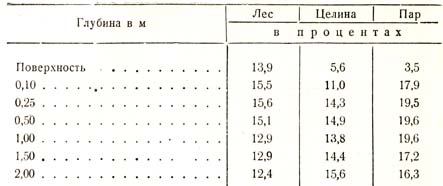
Эти же запасы влаги к концу вегетационного периода выраженные в миллиметрах, представляются в таком виде: под лесом – 456, под целиной – 473 и под паром – 641. На основании этих данных Г. Н. Высоцкий делает следующие заключения:
1. Поверхность почвы наиболее иссушается там, где она более открыта, т. е. на черном пару, затем под целиною и меньше всего под лесом.
2. Почва наиболее иссушается под целиною, затем под лесом: и менее всего под черным паром.
3. Подпочва, или грунт, иссушивается наиболее под лесом, затем под целиною и меньше всего под черным паром.
Исследования Близнина, Измаильского, Адамова, Тольского, Охлябинина, мои и других авторов, кроме указанных выше, а за границей Гоппе, Бюлера, Фрикке и др. впюлне подтверждают все вышеприведенные положения. Наиболее полная сводка всех русских наблюдений дана более других потрудившимся над данным вопросом Г. Н. Высоцким в его работе «О взаимных соотношениях между лесной растительностью и влагой», а также в труде П. В. Отоцкого «Грунтовые воды».
С лесоводственной точки зрения, однако, представляет большой интерес этот вопрос не только в его общей форме, но и в деталях, а именно: нельзя ли подметить какое-нибудь влияние на величину и распределение влаги в таких для нас существенных моментах, как те, которые характеризуют всякое насаждение, как-то: состав, форма, густота леса и пр. Многочисленные мои наблюдения над этого рода явлением дали мне возможность подметить четыре типа распределения влаги в почве, занятой лесом.
К первому надо отнести тот, вероятно, наиболее распространенный случай, когда некоторой мощности верхний горизонт почвы, прикрытый подстилкой, защищенный пологом и в то же время свободный от корней древесных растений, характеризуется большей влажностью по сравнению с такими же горизонтами безлесного пространства, причем в одних случаях в течение всего вегетационного периода, в других – только до осени. Первый случай наблюдался Раманном и Эбермайером, второй – мною, в сосновых жердняках. Для того чтобы представить себе, о чем идет речь, необходимо вспомнить, что влияние леса на влажность почвы, по верной характеристике Отоцкого, – это «алгебраическая сумма всех взаимно переплетающихся влияний», которые лес как живой организм производит на окружающую среду. В течение лета, в период сильного расходования влаги, верхний горизонт почвы под сосновым жердняком, будучи защищен от физического испарения пологом и подстилкой, физиологически не испаряет, потому что в нем нет деятельных древесных корней; в период же осеннего промачивания почвы он будет, по сравнению с таким же горизонтом полянной почвы, в менее выгодных условиях, так как будет получать меньше осадков, чем ничем не прикрытая почва; в летний период это обстоятельство также имеет место, но оно стушевывается перед другим моментом – сильного расходования влаги о поверхности открытой почвы и защиты от такого расхода почвы под насаждением. В этом первом типе более глубокие горизонты, пронизанные корнями, будучи весною одинаковой влажности, на песчаных, по крайней мере, почвах, как и соответствующие слои поляны, становятся в течение вегетационного периода суше. Наблюдая ход и распределение влажности почвы над и под сосновым жердняком, устанавливаем такую картину: сильная защитная способность такого густого и относительно невысокого полога соединяется в нем с сильной влагопотребляющей деятельностью; но здесь эти как бы борющиеся начала точно отмежевались в пространстве; слой почвы, в котором нет корней, находится под доминирующим влиянием защитной способности полога и подстилки, более глубокий горизонт, пронизанный корнями, наоборот, – под доминирующим влиянием влагопотребляющей деятельности; оттого один влажнее почти в течение всего года, соответствующих слоев поляны, другой наоборот – суше в течение всего года, кроме весны. Но указанная схема %не вполне применима к сосновому жердняку; в период осеннего промачивания почвы, как мы видели, открытая почва, находясь в более благоприятных условиях в отношении доступа осадков, становится теперь влажнее почвы под жердняком, где защитная способность полога, столь великолепно действовавшая во время лета, становится вместе с подстилкой отрицательным фактором, так как препятствует доступу осадков.
Если мы представим себе теперь не супеси и свежие супесчаные почвы, на которых мы наблюдаем наши сосновые жердняки, а более бедные и сухие песчаные почвы так называемого сухого бора, то встретимся с другой картиной распределения влаги. Здесь, в этом втором типе, даже верхние горизонты почвы, будучи равны весною по влажности с безлесным пространством, сохраняют это равновесие в течение всего вегетационного периода или становясь временами суше, но оно не носит постоянного характера, так как в периоды выпадения осадков почва открытых мест становится влажнее, а почва под пологом леса, таким образом, как бы суше; эти кривые как бы постоянно пересекаются. Этот случай представлен в природе спелыми сосновыми насаждениями так называемого сухого бора, где сильно развивается поверхностно стелющаяся корневая система, где почти нет подстилки и где защитная способность полога, благодаря малой сомкнутости этих насаждений, невелика.
Третий тип распределения влаги имеет много общего с только что описанным, так как в нем верхний, более влажный, горизонт почвы весьма ничтожен по своей мощности, иногда не более 5 см; все же остальные слои гораздо суше полянных и притом даже весною не уравниваются в количестве влаги с полянными. Этот случай представлен сложными насаждениями дуба с его спутниками, имеющими второй ярус из древесных пород, а третий из кустарников на суглинистых почвах; здесь густое древесное население, в особенности кустарниковое, развивает такую мощную корневую систему, что сильно высасывает влагу из почвы, благодаря чему почти на-нет сводится защитная способность трехъярусного полога. Этот тип распределения влаги я наблюдал в Шиповом лесу.
Наконец, к четвертому типу следует отнести насаждения, у которых почва и грунт весною влажнее соответствующих горизонтов безлесного пространства, и перевес этот как в верхних, так и нижних горизонтах сохраняется значительно дольше, чем во всех прежде рассмотренных случаях. Это – опушки, отдельные, лентообразные полосы леса, небольшие куртины леса среди степей, собирающие благодаря своему положению громадные количества снега.
К этим четырем случаям можно было бы прибавить еще пятый, наблюдаемый в лиственных насаждениях пойменных мест с близким уровнем грунтовых вод, где осенью, после опада листьев, грунтовая вода не потребляется больше древесного растительностью, поднимается выше и обусловливает большее увлажнение верхних горизонтов по сравнению с состоянием в них влажности в течение лета.
Исследования Гоппе в Мариабрунне, близ Вены, в 60-летнем буковом насаждении показали, что прореживание леса [1 Указание о влагонакопляющем действии полос и куртин леса в степях подчеркивает положительное значение полезащитного лесоразведения] увеличивает влажность почвы; на глубине 25 – 30 см в среднем в прореженном насаждении было больше влаги на 1,6%, чем в более сомкнутом, как благодаря большему проникновению осадков, так и в силу уменьшения числа деревьев, потребляющих влагу. Таким образом из этих и других подобных данных мы опять можем установить биологическую ценность как естественного, так и искусственного изреживания леса. С изреживанием насаждения не только будут входить под полог леса большие количества света, тепла, влаги, но и меньше потребляться последней, благодаря уменьшению числа потребителей, отчего подрост будет испытывать меньшую конкуренцию со стороны материнских растений. Что же касается непроизводительного расхода влаги путем физического испарения из почвы, то он не будет усиливаться, пока насаждение сохраняет свой характер, пока оно не лишилось мертвого покрова, пока не произошла перемена в составе живого покрова.
Теперь мы можем себе представить в более всестороннем развитии гидрологические условия жизни подроста. Чем отличается в этом отношении его жизнь от такого же подроста вне леса? Последний при сильном развитии засухи может пасть жертвой борьбы с нею, может умереть от недостатка влаги. Но может ли недостаток влаги послужить причиною отмирания самосева и подроста под пологом леса? Повидимому нет, так как здесь он защищен более влажной пеленой атмосферы, защищен от тепловых лучей и ветра; но на самом деле исследования проф. Раманна, отчасти Фрикке и мои, показывают иное. В сухих сосновых борах, например, самосев сосны в засушливое лето гибнет под пологом леса, гибнет не от недостатка света и не от других причин, а от недостатка влаги. Подрост на открытых пространствах имеет в своем распоряжении больший запас влаги в почве, чем подрост под пологом леса, но зато и внешние условия, окружающие первый, предъявляют к нему более высокие требования на транспирационую влагу; этих больших запасов влаги, которые мы наблюдаем на лесосеках, вообще на безлесных пространствах, может не хватить при большом физиологическом испарении подроста таких мест, да еще при конкуренции травянистого покрова, и растения умрут, побежденные засухою. Под пологом же леса результат может быть такой же, только от иных условий: здесь именно запрос на влагу, как это мы видели, невелик, но и влаги в почве тоже меньше, хотя и не в такой степени; подросту здесь приходится конкурировать с материнскими корнями, и весь вопрос в том, сумеет ли при данных условиях подрост свести свой водный баланс; он может его так же не свести здесь, как не сводит иногда и на открытых пространствах.
Когда мы рассматривали явление накопления влаги, то поставили вопрос, можно ли насаждения рассматривать как средство накопления влаги, и ответили на основании фактического материала на этот вопрос отрицательно; лес не накопляет влагу, но из накопленного запаса здесь ее больше поступает в почву и меньше непроизводительно стекает. Теперь перед нами аналогичный вопрос: можно ли рассматривать насаждения как средство сбережения влаги? Рассматривая часто насаждения с такой точки зрения, лесоводы иногда производят посадки под каким-нибудь защитным пологом леса. Лес, как мы видели, есть сильный потребитель влаги; древесные растения, за малым исключением, не принадлежат к типу сухолюбов, а наоборот – к растениям с большими запросами на влагу; этому способствуют также высоко поднятые над землею кроны, и лес поэтому и на основании всех эмпирических данных не есть сберегатель влаги, но вся организация его такова, что как в деле накопления, так и в деле расходования влаги непроизводительный элемент расход; сводится до минимума.[1 В таком же направлении трактует вопрос формула академика Г. Н. Высоцкого: лес сушит равнины и увлажняет горы. Эти утверждения по но вым материалам должны быть пересмотрены. Ред.]
В заключение уместно будет привести недавние исследования Бюлера, который изучал количество просачивающейся воды сквозь почву как открытых мест, так и закрытых лесом; для этого он в питомнике закапывал в почву громадной величины деревянные ящики, наполнял землею и одни засаживал древесными породами, другие оставлял голыми, третьи покрывал подстилкою или мхом и т. д.; с помощью особых сосудов улавливалось количество просачивающейся влаги как в указанных ящиках, так и почвах под пологом соседних естественных насаждений. За 21 месяц наблюдений просочилось следующее количество влаги, приведенное в табл. 36.
Таблица 36
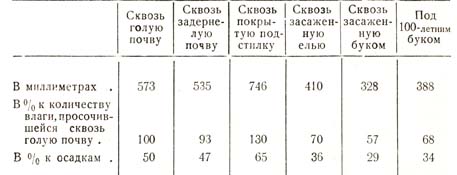
Здесь мы имеем как бы тот же опыт Эбермайера с тем различием, однако, что в ящики-лизиметры были помещены и корни древесных растений, и тогда сейчас же обнаружился другой результат, т. е. подтвердилась большая сухость почвы, занятой лесом, и благодаря этому меньшее количество просачивающейся влаги.
В заключение приведу сравнительные данные влажности почвы, занятой лесом, добытые мною при такого рода опыте: я наблюдал влажность почвы под трехъярусным дубовым насаждением Шилова леса на лесном суглинке на двух совершенно идентичных пробных площадях, причем первый год обе площади были под лесом, а затем лес на одной из них был вырублен, на другой оставлен; наблюдения же над влажностью продолжались и производились до глубины 4 м. Приводимые ниже данные представляют собою средние величины для всей четырехметровой толщи и представляют собою средние из нескольких определений в месяц и среднее из 20 – 40 данных, так как почвенные образцы брались с 10, 25, 50 см, а затем с каждого Чг м и притом каждый раз из двух параллельных скважин. Данные сосредоточены в табл. 37, которая комментариев не требует.
Таблица 37
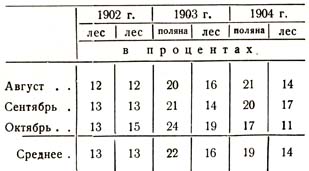
Резюмируя все сказанное, мы должны еще раз подчеркнуть, что обстановка леса под пологом несколько иная, чем та, которая обусловлена местным климатом и почвой, и что степень изменения занятой лесом среды будет различна в зависимости от состава леса, его возраста, формы, густоты и т. п.
ЛЕСНАЯ ПОДСТИЛКА
Деревья, соединяясь в насаждение, вносят еще один новый элемент, отсутствующий у деревьев, изолированно друг от друга растущих, хотя бы число их было несметное. К двум функциям листовых органов (дыхание и ассимиляция) насаждение присоединяет еще третью: листья после своего отпада дают в лесу так называемую лесную подстилку, иногда еще называемую лесным войлоком, а также мертвым покровом. Название подстилка объясняется тем, что она на самом деле в сельском хозяйстве служит подстилкою скоту в стойлах, заменяя собою солому; Насыщенная выделениями животных, она поступает затем в виде Удобрения в почву и потому играет большую роль в крестьянском хозяйстве. Не могу не обратить внимания на то, что она, в связи с условиями ее перегнивания, которые создаются самим насаждением (отенение, большая влажность, малый приток тепла), играет доминирующую роль в вопросе о влиянии леса на почву. Весь химизм лесных почв, насколько он обусловлен лесом, весь подзолообразовательный процесс коренятся главным образом в свойствах этой подстилки и условиях ее перегнивания; и если раньше старые лесоводы придавали ей громадное лесоводcnвенное значение, исходя из односторонних представлений о ее положительной роли и как источника непосредственного питания (до Либиха), и как мертвого покрова, защищающего почву от испарения влаги, то теперь, после всестороннего освещения ее роли, и пользы, и вреда ее, в зависимости от типа подстилки, – значение ее в жизни леса еще более возросло.
Количество подстилки зависит от соотношения между приходом ее, в силу отпада, и расходом, благодаря разложению. Оба момента, т. е. приход и расход, зависят в свою очередь от: 1) климата, 2) почвенных условий, 3) положения, 4) состава насаждений, 5) формы, 6) возраста и 7) густоты насаждения.
Чем больше условия местопроизрастания соответствуют данной породе, чем * выше бонитет почвы, – тем больше листвы падает на жердневый возраст и на насаждение средней полноты. Породы густо облиственные, теневыносливые, дают больше подстилки, чем светолюбивые со светлой кроной. Их можно распределить приблизительно в следующий убывающий ряд: пихта, ель, бук, граб, липа, клен, лещина, черная сосна, ольха, ильмовые, дуб, ясень, осина, сосна, береза и лиственница. Кроме того, состав насаждений влияет в том отношении, что листва разных пород отличается разной способностью к разложению, что зависит от нежной организации листовых органов, количества древесных частей в них, содержания дубильных веществ в них, смол и т. д. Имеет также значение, скручивается ли лист, когда ложится на землю, или нет; в последнем случае получается плотное, слегание, препятствующее доступу воздуха и способствующее тем образованию кислого перегноя.